| Головна |
| «« | ЗМІСТ | »» |
|---|
серотонін, або 5-гідрокси (5НТ), також є медіатором, що належать до групи біогенних амінів.
Серотонін утворюється з харчової амінокислоти - триптофану, що містить індольне кільце (рис. 9.14).
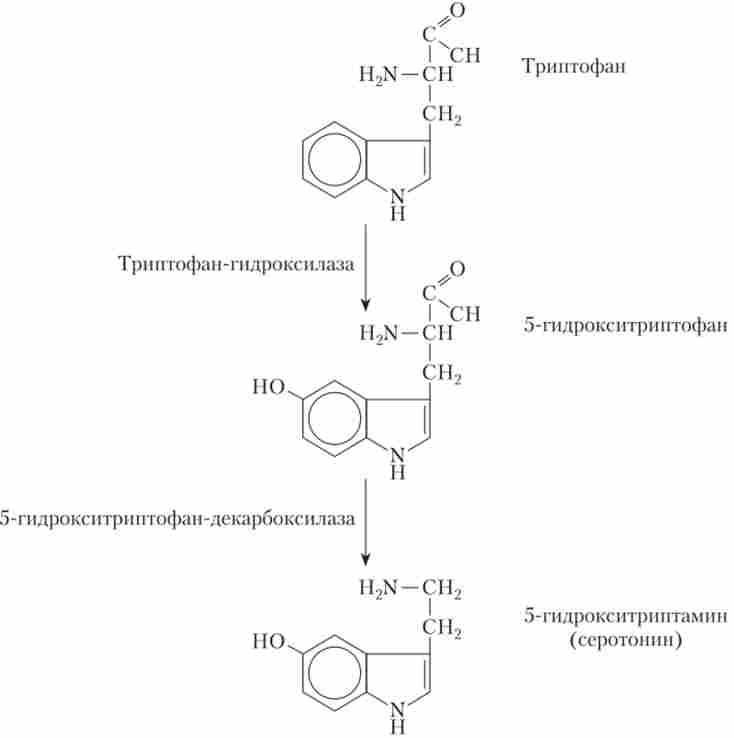
Мал. 9.14. Основний шлях біосинтезу серотоніну
Серотонін утворюється безпосередньо в закінченнях нейронів і депонується в везикулах. Будучи медіатором ЦНС, він досить широко представлений в периферичних системах, підвищуючи тонус гладкої мускулатури і звужуючи просвіт судин. Збільшуючи проникність капілярів, 5НТ може викликати набряки. Сприяючи звуження артерій мозку, серотонін може бути причиною нападів мігрені. Присутня в тромбоцитах крові, серотонін звужує судину в місці його пошкодження і прискорює утворення в цьому місці тромбу.
В ЦНС нейрони, що містять серотонін, розташовані компактно, уздовж середньої лінії довгастого і середнього мозку - в ядрі шва (див. Рис. 9.10; рис. 9.15).
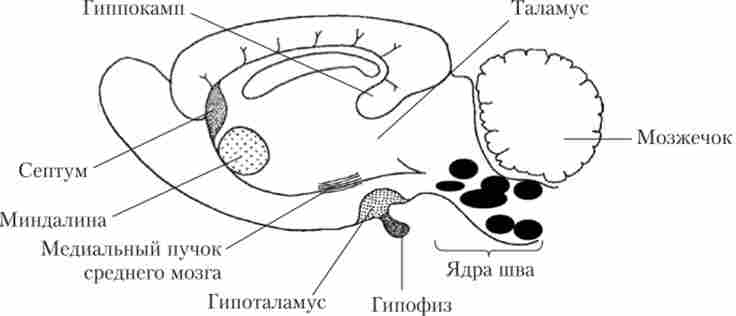
Мал. 9.15. Серотонінергічні ядра шва і структури мозку, в які проектуються серотонинергичні волокна
Найбільший тракт, в закінченнях аксонів якого виділяється 5НТ, - медіальний пучок середнього мозку. Причому клітини, розташовані в спинному ядрі шва, дають аксони в базальні ядра і мозочок, а в медіальному ядрі шва - в гіпокамп і перегородку. Велике число 5НТ-ергічних закінчень виявлено в структурах кори великих півкуль. У задній частині ядер шва, розташованих уже в довгастому мозку, виявлена група серотопііергічних нейронів, що дають спадні шляху в стовбур і СМ. Ці нейрони мають велике значення для регуляції вегетативних реакцій, рухової активності, модуляції больових відчуттів.
Серотонін є одним з найбільш поліфункціональних медіаторів, беручи участь в регуляції емоційного стану і будучи одним з факторів в системі внутрішнього підкріплення; він впливає на цикл «сон - неспання», контролює роботу сенсорних систем; крім того, він пов'язаний з навчанням, особливо на тлі позитивного підкріплення, і впливає на рухову активність. Іншими словами, 5НТ-система пов'язана з усіма аспектами діяльності мозку, тому не дивно, що рецептори до серотоніну відрізняються великою різноманітністю.
В даний час налічується сім класів 5НТ-рецепторів. 5НТГ клас поділяється на вісім підкласів, локалізованих в межах
ГМ. Всі рецептори першого класу пригнічують активність АЦ в клітці через Gy-білки, що в підсумку призводить до гіперполяризації мембрани. Активація рецепторів класу 5НТ{ на рівні цілого організму призводить до зниження агресивності, тривожності, підвищення рухової активності і стимуляції статевої поведінки.
рецептори 5НТ2-класу поділяються на три підкласу (Л, В, С) і локалізовані вони в ГМ. Механізм їх дії пов'язаний з G ^ -белкамі, активацією фосфоліпази З і стимуляцією фосфоінозітольной системи. Саме з впливом на 5НТ2А-рецептори пов'язують потужні психотропні ефекти галлюциногена LSD. Активація 5НТ2-рецеіторов, і особливо 5НТ2С, викликає почуття заспокоєності, зниження спраги і апетиту.
рецептори 5НТ3-класу принципово відрізняються від всіх інших рецепторів до серотоніну, оскільки вони не є метаботропнимі, а являють собою серотонін-активуються катіонні канали, відкриття яких призводить до деполяризації клітини. Розташовані рецептори третього типу в корі, лімбічної системи, стовбурі. рецептори 5НТ3-класу контролюють викид інших медіаторів, як збудливих, так і гальмівних. На рівні цілого організму вплив агоністів серотоніну на ці рецептори викликає нудоту, пригніченість, погіршує навчання, пригнічуючи довгострокової потенциации в гіпокампі.
Рецептори класів 5НТ4, 5НТ6, 5НТ7, виявляються в корі великих півкуль, лімбічної системи, таламусі, гіпоталамусі, стриатуме, об'єднуються тим, що через С5-білки підвищують активність АХ, активуючи клітку. агоністи 5НТ7 беруть участь в регуляції циркадних ритмів і синхронізації сну зі світловим циклом. Найменше відомостей є про рецепторах класу 5НТ5.
У синаптичної щілини частина 5НТ руйнується, а частина, після взаємодії з рецепторами, захоплюється молекулами-транспортерами і переноситься назад у пресінаітічне закінчення для повторного використання. Є відомості про те, що частина серотоніну, як і інших біогенних амінів, захоплюється з синаптичної щілини в гліальні клітини - астроцити, а вже з них - в закінчення нейрона.
Вплив серотониновой системи на сприйняття навколишнього середовища дуже велике. Описано серотонинергичні шляху до тих нейронам сенсорних зон кори великих півкуль, які обробляють тактильну, слухову і зорову інформацію. Ці серотонинергичні впливу, переважно гальмівні, перешкоджають зайвій поширенню сигналів по сенсорним полях, як би фокусуючи їх образи, роблячи контрастніше і чіткіше. Подібне дія серотонинергичні впливу надають і на асоціативні зони кори, підтримуючи їх роботу в оптимальному режимі, перешкоджаючи зайвому порушення.
Серотонінергічні проекції гальмують больові відчуття, що можна довести, дратуючи в експерименті ядра шва. Порушення передається на гальмівні вставні нейрони задніх рогів СМ, які гальмують активність нейронів, що входять до складу больових шляхів.
Вплив серотоніну на центри мотивацій і емоцій позначається в зниженні біологічних потреб, зменшення апетиту, спраги, зниження тривожності.
Маючи пряме відношення до систем позитивного і негативного підкріплення, серотонинергична система бере участь в процесах навчання. Зниження активності 5НТ-системи мозку призводить до цілого ряду розладів: депресії, підвищення тривожності, страхів, безсоння. Тому робляться постійні спроби створення лікарських засобів на базі аналогів 5НТ.
агоністи 5НТ2-рецепторів використовують для зняття нападів мігрені. Навпаки, блокатори 5НТ2-рецепторів прописують при підвищеному артеріальному тиску і зайвій вазі для зниження апетиту. Виборчий інгібітор зворотного серотонінового транспортера - флуоксе- тин - є хорошим антидепресантом.