| Головна |
| «« | ЗМІСТ | »» |
|---|
дофамін - катехоламін, що є попередником НА в загальній для катехоламінів ланцюжку біосинтезу (див. рис. 9.11). Дофамінергічні нейрони не містять фермент дофамін-р-гідроксилази, і тому в них не синтезується НА. Дофамін зустрічається в організмі в якості медіатора майже виключно в межах ЦНС.
Дофамінсодержащіе нейрони зібрані в компактні структури в трьох відділах ГМ: чорної субстанції середнього мозку, покришці середнього мозку і деяких ядрах гіпоталамуса. Нейронів, що містять Д, відносно небагато. Так, у чорній субстанції середнього мозку таких нейронів близько 7 тис. Але зате кожен з цих нейронів утворює приблизно по 250 тис. Синаптичних закінчень на нейронах переднього мозку. Аксони дофамінергічних нейронів чорної субстанції направляють свої аксони в хвостате і чечевицеподібних ядро базальних гангліїв, які разом називаються стриатум, або смугасте тіло (рис. 9.12).
Цей шлях називається нігростріарной. Функції цієї системи полягають у підтримці загального рівня рухової активності, фіксації рухових програм, забезпеченні точності довільних рухів. Аксони дофамінергічних нейронів покришки направляються до структур лімбічної системи (мигдалеподібного комплексу, гиппокампу), нюхової, асоціативної лобової, премоторної, моторної, зорової корі. Дофаминергична іннервація цих структур через мезолімбіче- ські шляху доводить участь Д в підтримці тонусу вищих центрів сенсорного сприйняття, когнітивних функцій, емоційного стану.
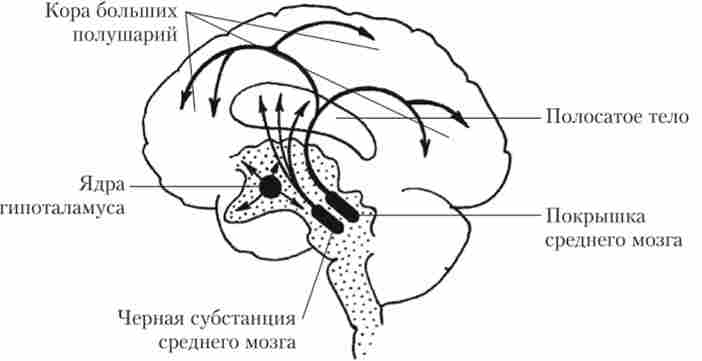
Мал. 9.12. Схема розташування в головному мозку нейронів, що виробляють дофамін, і розподіл їх аксонів
Гіпоталамічні дофаминергичні нейрони мають короткі аксони і входять в систему регуляції нейроендокринних зон і потребностно- мотиваційних центрів гіпоталамуса, забезпечуючи секрецію либеринов і статинів нейроендокринними клітинами (див. Рис. 9.12).
Дофамін синтезується головним чином в закінченнях нейронів і депонується в везикулах. Механізм викиду цього медіатора в щілину не відрізняється від такого для інших біогенних амінів. У синаптичної щілини Д досягає рецепторів на пресинаптичної мембрани.
Всі рецептори до Д є повільними метаботропнимі і змінюють активність АЦ через G-білки. Виділяють п'ять підкласів рецепторів до Д, що розрізняються по спорідненості до Д і його агонистам, антагоністів, локалізації в мозку, розташуванню на нейронах, спрямованості впливу на АЦ (рис. 9.13).
Більшість рецепторів до Д відносяться до підкласу і розташовуються на постсинаптичних мембранах, активуючи АЦ і, таким чином, і клітку шляхом підйому рівня цАМФ. найбільше JX-рецепторів на клітинах стрі- Атума, нової кори, лімбічної системи. Серед рецепторів підкласу Д! особливо виділяють рецептори Д5, які відрізняються дуже високою чутливістю до Д і локалізовані на клітинах лімбічної системи.
Рецептори підкласу Д2, на противагу Д1? пригнічують АЦ через С, -белок. Рецепторів цього підкласу набагато менше, ніж Д1? локалізовані вони в блідій кулі, таламусі, нової корі, лімбічної системи. У підкласі Д2 особливо виділяють рецептори Д3 і Д4, причому для рецепторів Д3 частіше характерна пресинаптична локалізація (див. рис. 9.13), і, по-видимому, вони покликані послабляти ефекти занадто сильною активації дофаминергичної системи. рецептори Д2-нодкласса проявляють особливу чутливість до нейролептиків - головному класу ліків, що володіють антипсихотичним дією, застосовуваним при лікуванні гострих психозів, шизофренії, білої гарячки.
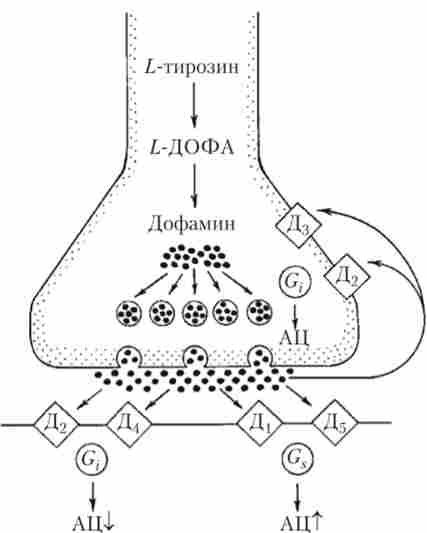
Мал. 9.13. Схема розташування дофамінових рецепторів різних підкласів
Після впливу на рецептори Д на 80% захоплюється специфічним транспортером, який повертає його в пресіпаптічне закінчення, де медіатор знову закачується в везикули або руйнується КОМТ або МАО.
Особливе значення Д має в зв'язку з тим, що він єдиний серед всіх медіаторів є безпосереднім регулятором ендокринної системи, виконуючи функції пролактостатіна. Пролактин - гормон передньої долі головної ендокринної залози нашого організму - гіпофіза. Пролактин стимулює виділення молока молочними залозами, одночасно блокуючи виділення ряду статевих гормонів гіпоталамусом і гіпофізом. Таким чином, менструальний цикл у жінки, що годує пригнічений, і ймовірність завагітніти знижена, хоча не до нуля. В потрібний час Д, виділяючись в гіпоталамусі, зменшує секрецію пролактину і припиняє лактацію (виділення молока), відновлюючи менструальний цикл і здатність жінки до нової вагітності.
Підводячи підсумок відомостями про Д як про один з головних регуляторних факторів мозку людини і тварин, потрібно сказати про те, що Д є одним з внутрішніх факторів винагороди, виділяючись в тих випадках, коли людина досягає поставленої мети, задовольняючи свої потреби і досягаючи поставлених цілей. Беручи участь в генезі позитивних емоцій, дофаминовая система знижує рівень біологічних потреб: статевий, харчовий, приводячи до виникнення позитивних емоцій, почуттю задоволення.
Інша найважливіша роль дофаминовой системи - регуляція рухової активності, яка ініціює зміни режиму локомоции (нерухомість - ходьба, ходьба - біг, біг - зупинка і. Т. П.). Крім того, дофаминовая система необхідна для виконання рухових програм і забезпечення точності довільних рухів.
Нарешті, Д є важливим ендокринним регулятором, визначаючи режим роботи репродуктивної системи.