| Головна |
| «« | ЗМІСТ | »» |
|---|
дуплікації, в строгому сенсі цього слова, є дворазове повторення одного і того ж ділянки хромосоми. Відомі випадки багаторазових повторень, або мультипликаций, якої-небудь ділянки. Їх називають також ампліфікації.
Дуплікації можуть відбуватися в межах однієї і тієї ж хромосоми або супроводжуватися перенесенням копії ділянки генетичного матеріалу на іншу хромосому. Повтори, що виникли в одній хромосомі, можуть розташовуватися тандемно (ABCBCDE...) або інвертовану (ABCCBDE...). розрізняють також термінальні повтори, якщо Дуплікація зачіпає кінець хромосоми.
Мабуть, головною причиною множинних повторів ділянок генетичного матеріалу є так званий нерівний кросинговер. Найбільш відомий приклад участі цього механізму в генерації повторів отримано для локусу ваг (В) у D. melanogaster. домінантна мутація В (Полосковідние очі) являє собою дуплікацію невеликого сегмента (16А) у кінця Л'-хромосоми (на 57,0 сМ від кінця хромосоми) (рис. 9.3). У лінії, гомозиготною по ваг, зрідка з частотою близько 6 х Ю ^1 на гамету спостерігаються реверсії - вищепленію мух дикого типу В *, а також мух з ще вужчими очима, ніж у В. Цей посилений варіант ваг названий ультра-йог і позначається ВВ. У мух В В сегмент 16А в Х-хромосомі виявився вже тріпліцірованним, в той час як у ревертанти В ' той же сегмент представлений в однині.
Механізм цього прояву пояснив А. Стертевант ще в 1925 р на основі генетичного аналізу. Ділянки зліва і праворуч від ваг були марковані тісно зчепленими мутаціями: / - forked (56,7); В - ваг (57,0); fu - fused (59,5). Цифри в дужках - точки локалізації (сМ). При схрещуванні:
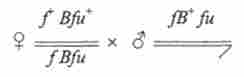
поряд з усіма очікуваними класами розщеплення з'являлися також мухи В ', нормальні за формою очі, і ВВ - ультра-йаг. Одні й другі були одночасно рекомбінантними по генам / і/ І. Ці незвичайні сегреган- ти виникали тільки в результаті мейозу у самок В / В. У реципрокною комбінації $ В * / В " х В вони не були отримані.
На підставі отриманих результатів А. Стертевант припустив, що мухи ультра-йаг і Ваг '- реципрокні продукти нерівного кроссінго- віра між дупліціроваться ділянками Х-хромосоми.
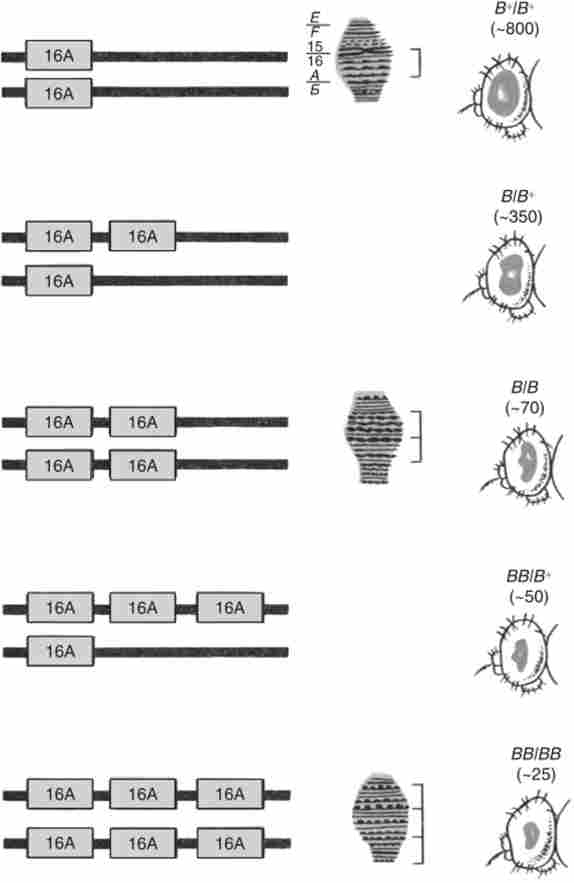
Мал. 9.3. Фенотипічніпрояв повторів одного і того ж ділянки (16А) в Л'-хромосомі О. melanogaster - зміна ознаки Ваг.
У дужках - середнє число фасеток в очах мух відповідного генотипу
Пізніше на матеріалі гігантських хромосом було показано, що ул'тра- ваг - дійсно тричі повторений сегмента 16А, дупліціроваться у мутанта і одиничного у дикого типу ваг+.
Мабуть, і вихідна «мутація» ваг виникла в результаті нерівного кросинговеру (рис. 9.4). В цьому випадку дуплікації і делеции можуть мати загальний механізм виникнення.
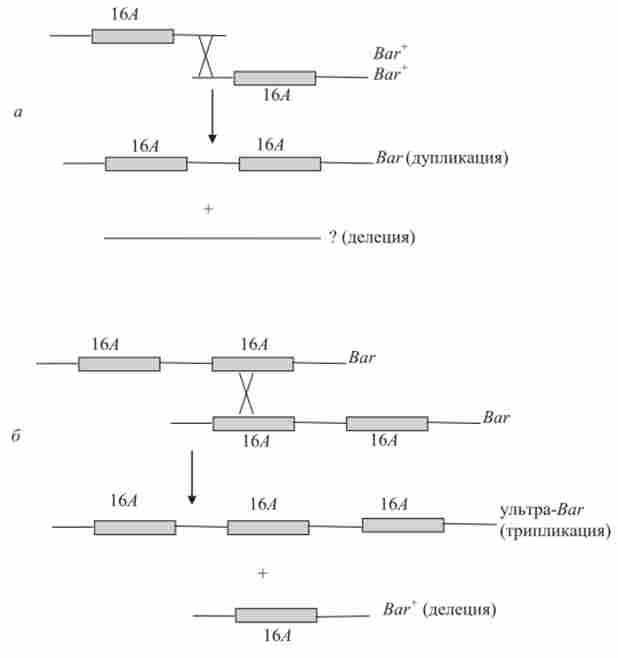
Мал. 9.4. Можливий механізм виникнення дуплікацій і поділів в результаті нерівного кросинговеру на прикладі мутацій ваг в. ^ - хромосомі D. melanogaster: а - при виникненні мутації Ваг. б - при виникненні мутації ультра-itor; 16А - найменування диска, дупліціруемого в політенних хромосомах
Дуплікації і делеции можуть виникати в результаті нерівного кросинговеру між сестринськими хроматидами або навіть в результаті рекомбінації в межах однієї і тієї ж хроматиди.
Фенотипічніпрояв дуплікацій показано на прикладі ряду генів у дрозофіли, як рецесивних: white (W), scute (sc), achaete (ас) - редукція щетинок, vermilion (V) - яскраво-червоні очі, так і домінантних: Ваг (В), Hairy wing (Hw) - волохаті крила.
Цитологічних гетерозиготи по дуплікацій виявляються схоже з гетерозиготами по ділок - утворюється петля при кон'югації хромосом.
Дуплікації і делеции часто виникають в результаті розривів хромосом, що викликаються різними повреждающими агентами, іонізуючою радіацією, хімічними мутагенами, вірусами та ін., А також факторами рекомбінації у самців О. melanogaster (МЯ), що представляють компоненти системи гібридного дисгенезу. Крім того, дуплікації і делеции можуть бути результатом кросинговеру у гетерозигот по інверсія і транслокаціях.
Дуплікації і інші повтори зазвичай не надають такого негативного впливу на життєздатність, як делеції і дефішенсі. Подібні елементи часто повторюються в геномах різних організмів. Так, К. Бріджес звернув увагу на подібний малюнок розташування дисків в двох однаково оріенгірованних ділянках П хромосоми дрозофіли. Мабуть, ці ділянки являють собою прямий повтор. На гігантських хромосомах дрозофіли також виявляються інвертовані повтори. На існування повторів в геномах інших організмів вказує, наприклад, спаровування окремих ділянок хромосом при мейозі у гаплоїдних рослин, які можуть бути отримані експериментальним шляхом. За відсутності гомологів у гаплоидов спостерігаються ділянки синапсиса. Це так зване гомеологічне спарювання.
При дуплікації домінантного алеля будь-якого гена, що супроводжується транспозицией копії на іншу хромосому, змінюється і картина розщеплення, якщо схрещування проводять з рецесивним мутантом по тому ж гену. Замість моногибридного розщеплення 3: 1 у диплоїдних організмів спостерігається полімерне розщеплення 15: 1. Можливість клонування генів, перевірка гібридизації їх ДНК і визначення первинної структури генів часто розкривають високий рівень гомології полімерних генів. Наприклад, два гени, що кодують ізозімов (т. Е. Ферменти з однаковою функцією) кислої фосфатази у дріжджів ЯассІ. сегеутае, - РКО 3 і р1ю 5 - гомологични на 80%. Очевидно, ці гени колись виникли в результаті дуплікації.
Як показав І. А. Рапопорт для локусу ваг, експериментальне повторення одного і того ж гена (до 8 разів) може досягатися за рахунок багаторазових дуплікацій. Багаторазові повтори - ампліфікації - виявлені в культурах клітин ссавців, стійких до ряду агентів, що ушкоджують: метатрексат, колхицину, іонів важких металів та ін. В цих випадках кратність повторення одного і того ж ділянки генетичного матеріалу, що включає, наприклад, ген дигідрофолатредуктази, що зв'язує метатрексат, може досягати декількох сотень і навіть більше тисячі. У присутності метатрексата як селектірующего агента формуються цілі хромосоми з центромерами, що містять один і той же повторюваний фрагмент ДНК розміром в 30 000-40 000 п. Н. При цьому стійкість до метатрексат може підвищуватися до 100 тис. Разів. У процесі такої ампліфікації бере участь механізм рекомбінації.
У дріжджів аналогічно амплифицируют ділянку, що містить ген CUP, що обумовлює стійкість клітин до іонів міді. За відсутності селективного агента ці відібрані генетичні конструкції виявляються нестабільними, і стійкість швидко втрачається.
Багаторазові повторення одного і того ж ділянки характерні для генів рибосомной ДНК у дрозофіли. Близько 130 повторів цих генів зібрано в локусі bb {bobbed), який присутній як в Х-, так і в У-хромосомі цієї комахи. мутанти bb несуть делеции генів РДНК. Протягом декількох поколінь у мух, гомозиготних по делеции здебільшого РДНК, відбувається відновлення числа генних копій, характерних для дикого типу. Мультиплікація, або ампліфікація РДНК, супроводжується підвищенням частоти кросинговеру між X- і У-хромосомами в районі bb.
Дуплікації грають істотну роль в еволюції геному, оскільки вони створюють додаткові ділянки генетичного матеріалу, функція яких може бути змінена в результаті мутацій і подальшого природного відбору.