| Головна |
| «« | ЗМІСТ | »» |
|---|
Зміна чергування генів в хромосомі в результаті інверсії - тип перебудов, що найчастіше зустрічаються в природних популяціях. Залежно від розташування кінців (кордонів) перебудови по відношенню до центромере інверсії ділять на періцентрічні, захоплюючі центромеру і включають її в інвертований ділянку, і парацентричної, що не включають центромеру в інвертований ділянку.
Інверсія - це широко поширений шлях еволюційного перетворення генетичного матеріалу. Наприклад, людина і шимпанзе відрізняються за кількістю хромосом: у людини 2п = 46, а у шимпанзе 2п = 48. Хромосома 2 людини містить велику частину матеріалу, гомологичного додаткової парі хромосом шимпанзе. Крім того, відмінності стосуються чотирьох хромосом: 4,5,12 і 17-й, - в яких відбулися періцентрічні інверсії.
Інверсія призводить до зміни зчеплення генів, інший їх лінійної послідовності, ніж у вихідної форми. Цей ефект можна виявити, якщо інверсія в гомозигот нелегальний. Рецессивная летальність часто супроводжує інверсія як результат локалізації точок розривів в життєво важливих генах або як наслідок ефекту положення.
Інший важливий наслідок інверсії - придушення кросинговеру, якщо інверсія знаходиться в гетерозиготі. Це властивість інверсій широко використовують при створенні збалансованих ліній, гетерозиготних по летальним мутацій і не руйнуються кросинговером по потрібної хромосомі. У гетерозигот по інверсія кроссинговер не пригнічуючи, але наслідком його є утворення нежиттєздатних суперечка у рослин або зигот у тварин.
У гетерозигот по інверсія на цитологічних препаратах виявляють характерні петлі - результат кон'югації структурно зміненої і нормальної хромосом. Якщо в такий петлі, т. Е. В інвертованому ділянці, відбудеться одиночний кроссинговер, то в разі парацентричної інверсії виникає одна хроматида з двома центромерами, які її порвуть при розходженні в анафазе. Утворений бесцентромерний фрагмент також буде втрачено. В результаті з чотирьох гамет повноцінними будуть тільки дві. Тільки вони здатні при заплідненні дати життєздатні зиготи. При гетерозиготності по періцентрічної інверсії кроссинговер не перешкоджає нормальному розбіжності всіх хроматид. Проте повноцінними знову будуть тільки два продукти мейозу з чотирьох, оскільки дві хроматиди несуть делеции деяких генів.
У самок дрозофіли аномальні хромосоми, що утворилися в результаті кросинговеру в гетерозиготних інверсіях, зазвичай відходять в полярне тільце, тому гетерозиготность по інверсія не позначається на їх фертильності. У багатьох організмів гетерозиготи по довгих інверсія виявляються полустерільнимі внаслідок кросинговеру.
Ми розглянули несприятливий ефект 'одиночного кросинговеру. У той же час подвійний кросинговер у гетерозигот по інверсії може призводити до утворення цілком життєздатних гамет. Конверсія в межах гетерозиготной інверсії відбувається нормально, якщо вона не пов'язана з реципрокною рекомбинацией, як показав А. Човник для локусу гу (Темно-червоні очі) І. melanogaster.
Хромосома може нести не тільки одну інверсію, але дві непере- кривает і дві, що перекриваються повністю або частково. Гетерозиготність по таким складним перебудов також ідентифікується цитологічних за характером кон'югації хромосом (рис. 9.5).
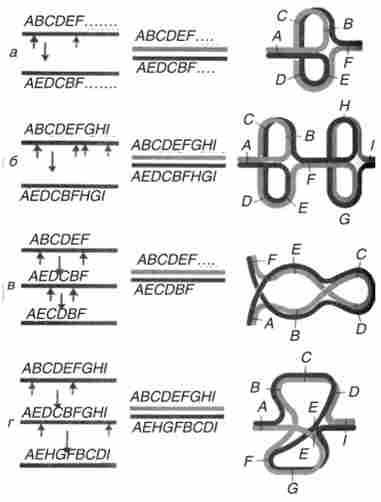
Мал. 9.5. Характер кон'югації хромосом при гстерозіготності: а - по одній інверсії; б - за двома неперскривающімся інверсія; в - по одній інверсії в межах іншої; г - за двома частково перекриваються інверсія. Короткі стрілки зліва - точки розривів при утворенні інверсії
Порівняння гігантських хромосом роду Drosophila показує, що дивергенція їх видів супроводжувалася складними перекриваються і не перекриваються інверсіями. Так, при вивченні кон'югації хромосом у гібридів D. virilis і D. littoralis Н. Н. Соколов виявив, що хромосоми цих видів розрізняються шістьма інверсіями.
Таким чином, з огляду на наслідки гетерозиготних інверсій для мейозу, слід підкреслити, що вони можуть служити факторами ізоляції і сприяти еволюційної дивергенції нових форм, що утворюються в межах виду.