
| Головна |
| «« | ЗМІСТ | »» |
|---|
Катаболізм вищих жирних кислот здійснюється переважно шляхом окислення. Ненасичені вищі жирні кислоти попередньо відновлюються до граничних кислот. Граничні (насичені) жирні кислоти окислюються поступово, шляхом відщеплення від їх молекул ацільних 2-вуглецевих фрагментів СН3-СО (рис. 9.15). Розпад вищих жирних кислот проходить в кілька стадій. Всі стадії окислення прискорюються специфічними ферментами.
Початкова стадія катаболізму жирних кислот полягає в активації. Вільна жирна кислота незалежно від довжини вуглеводневого ланцюга хімічно інертна і не може бути піддана біохімічним перетворенням, поки не буде активована. Активація жирної кислоти R-COOH протікає на зовнішній поверхні мембрани мітохондрій за участю АТР, коферменту-А (HS-KoA) і іонів Mg2'. Реакція каталізується ферментом ацил-КоА-синтетазой:
В результаті утворюється ацил-КоА (R-CO-S-KoA) - активна коензімную форма кислоти.
Подальше окислення жирних кислот відбувається на внутрішній стороні мембрани мітохондрій.
Коензімную форми кислот ацил-КоА, як і вільні жирні кислоти, які не мають здатність проникати всередину мітохондрій. З цитоплазми до внутрішньої сторони мембрани мітохондрій ацил-КоА переноситься карнітином. Спочатку карнітин реагує з ацил-КоА:
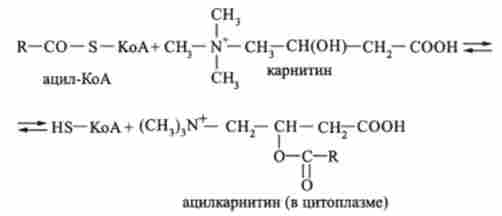
В результаті в цитоплазмі утворюється ацилкарнітину. який проходить через мембрану всередину мітохондрії, де розпадається, вивільняючи ацил-КоА:
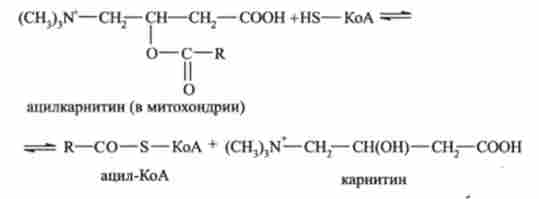
Окислення жирних кислот всередині мітохондрій протікає через кілька послідовних стадій.
Перша стадія. Ацил-КоА реагує з FAD і піддається ферментативному отщеплению водню - дегидрированию. При цьому ацил-КоА втрачає 2 атома водню (окислюється), перетворюючись в коа-ефір ненасиченої кислоти еноіл-КоА, з'єднання з подвійним зв'язком:
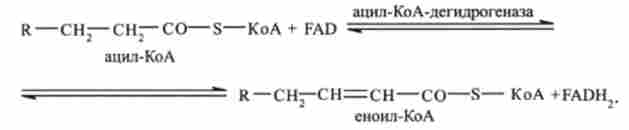
Слід помститися, що існує кілька ацил-КоА-дегідрогеназ, кожна з яких має специфічністю по відношенню до ацил-КоА з певною довжиною вуглецевого ланцюга.
Друга стадія. Еноіл-КоА (за участю ферменту еноіл-КоА-гідратази) приєднує молекулу води. Відбувається гідратація подвійного зв'язку. В результаті утворюється р-оксіаціл-КоА:
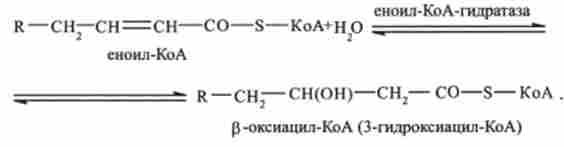
Третя стадія. р-оксіаціл-КоА дегидрирующей з утворенням 3-оксіаціл- КоА (каталізується ферментом N А Незалежної дегідрогеназ):
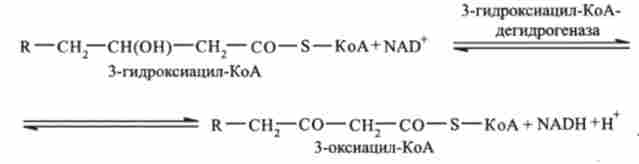
четверта стадія - тіолазная реакція. В ході тіолазной реакції відбувається розщеплення 3-оксіаціл-КоА з утворенням укороченою на два вуглецевих атома молекули ацил-КоА і ацильного 2-вуглецевого фрагмента СН3-СО, пов'язаного в ацетил-КоА (реакція каталізується ацетил-КоА-ацілтрансферази):
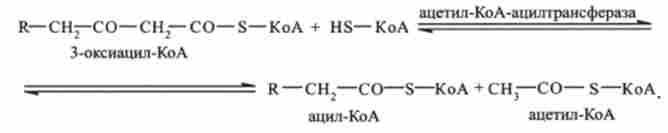
Утворився ацетил-КоА включається в цикл трикарбонових кислот. Скорочений на два вуглецевих атома ацил-КоА знову проходить весь шлях Р-окислення аж до утворення 4-вуглецевого з'єднання бутіріл-КоА. Бу- Тіріл-КоА потім окислюється до 2 молекул ацетил-КоА, які також включаються в цикл трикарбонових кислот. На цій стадії катаболізм жирної кислоти завершується.
У кожному циклі розщеплення жирних кислот утворюється р-кетоацил-КоА з усе більш укороченою вуглеводневої ланцюгом, тому розглянутий процес в цілому можна називати Р-окисленням жирних кислот. NADH і FADH2, що утворюються при Р-окисленні, потім віддають свої електрони мітохондріального дихального ланцюга.
При окисленні жирної кислоти, що містить п вуглецевих атомів, відбувається (л / 2-1) цикл Р-окислення. Наприклад, при окисленні пальмітинової кислоти (С |6) Повторюється 7 циклів р-окислення. Отже, сумарне рівняння р-окислення активованої пальмітинової кислоти має вигляд
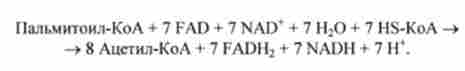
Молекула пальмітинової кислоти перетворюється в 8 молекул ацетил-КоА і додатково 7 молекул NADH і 7 молекул FADH2. Потім при окисленні NADH синтезуються 2,5 молекули АТР, а при окисленні FADH2 - 1,5 молекули АТР. Таким чином, окислення 1 молекули пальмітинової кислоти призводить до синтезу 106 молекул АТР з ADP і Р, (з урахуванням витрати 2 молекул АТР на освіту 1 молекули пальмитоил-КоА і синтезу 1 молекули GTP на кожну молекулу аце- тил-КоА в циклі лимонної кислоти ). Це означає, що енергетичний вихід біоокислення пальмітинової кислоти становить 33%.
Слід зазначити, що якщо остаточним продуктом р-окислення вищих жирних кислот з парним числом вуглецевих атомів є ацетил-КоА, то з непарним - пропіоніл-КоА.
Окислення ненасичених жирних кислот, в принципі, відбувається так само, як і окислення насичених, але має ряд особливостей. До подвійного зв'язку ланцюг такий кислоти коротшає в результаті звичайного р-окислення з утворенням / * мс-еноіл-КоА. Далі фермент ізомерази відсуває подвійну зв'язок і утворює трансізомери активованої кислоти т /? анс-еноіл-КоА:
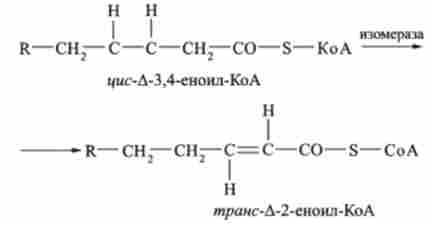
При р-окисленні жирних кислот, що мають дві і більше ненасичених зв'язку, потрібно ще один додатковий фермент.
Природні масла містять головним чином жирні кислоти з парним числом вуглецевих атомів. Однак в оліях багатьох рослин і деяких морських організмів присутні жирні кислоти з непарним числом атомів вуглецю. Крім того, у жуйних тварин при перетравленні вуглеводів в рубці утворюється велика кількість пропіонової кислоти СН3СН2СООН з трьома вуглецевими атомами. Пропионат всмоктується в кров і окислюється в печінці та інших тканинах.
Жирні кислоти з непарним числом вуглецевих атомів окислюються таким же чином, як і жирні кислоти з парним числом, з тією лише різницею, що на останньому етапі розщеплення (р-окислення) утворюється одна молекула пропіоніл- КоА і одна молекула ацетил-КоА, а не дві молекули ацетил-КоА.
Активоване 3-вуглецевий фрагмент - пропіоніл-КоА - включається в цикл трикарбонових кислот після перетворення в сукцинил-КоА.
В реакції ізомеризації-КоА бере участь кофермент дезоксі- аденозилкобаламін, похідне вітаміну В12.
Пропионат утворюється також при розщепленні чотирьох амінокислот - валіну, ізолейцину, метіоніну, треоніну - і з холестерину.