| Головна |
| «« | ЗМІСТ | »» |
|---|
У біосинтезі ліпідів de novo (ли погенез) використовується значна частина жирних кислот, моногліцеридів і гліцерин, який звільняється при гідролізі жирів, що надходять з їжею. Це пов'язано з тим, що ліпіди кожного організму, так само як білки і вуглеводи, мають індивідуальний склад і будова.
Синтез жирних кислот протікає в цитоплазмі клітини. В мітохондріях в основному відбувається подовження ланцюгів жирних кислот (рис. 9.16).
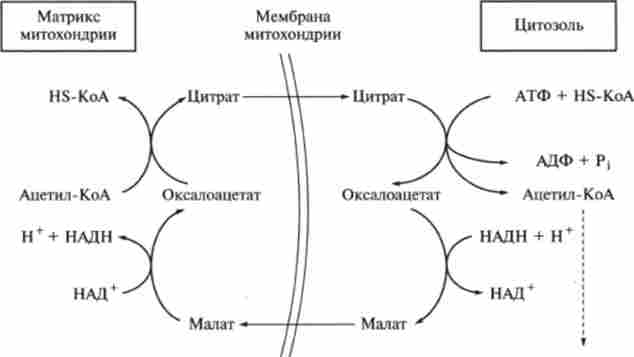
Мал. 9.16. Взаємодія мітохондріальної і внемітохондріальной систем біосинтезу
de novo жирних кислот
Встановлено, що в цитоплазмі печінкових клітин синтезується пальмітинова кислота (16 вуглецевих атомів). З неї або з жирних кислот екзогенного походження (т. Е. Що надходять з кишечника) в мітохондріях печінкових клітин утворюються жирні кислоти, що містять 18, 20 і 22 вуглецевих атома.
Мітохондріальна система біосинтезу жирних кислот здійснює тільки подовження існуючих в організмі среднецепочечних жирних кислот. Повний біосинтез пальмітинової кислоти з ацетил-КоА активно протікає поза мітохондрій в цитоплазмі іншим шляхом.
У немітохондріальних система біосинтезу de novo жирних кислот знаходиться в розчинній (цитозольних) фракції клітин багатьох органів, зокрема печінки, нирок, мозку, легенів, молочних залоз, а також в жировій тканині. Біосинтез жирних кислот протікає за участю NADPH, АТР (на малюнку в російській написанні НАДФН, АТФ і т. Д.), Іонів марганцю Мп2+ і бікарбонату НСОз- (В якості джерела СО2); субстратом є ацетил-КоА, кінцевим продуктом - пальмітинова кислота.
Будівельним блоком для синтезу жирних кислот в цитозолі клітини служить ацетил-КоА, який в основному надходить з мітохондрій. Спочатку внутрімі- тохондріальний ацетил-КоА взаємодіє з оксалоацетата, в результаті чого утворюється цитрат. Реакція каталізується ферментом цитрат і нтазой. Утворився цитрат переноситься через мембрану мітохондрій в цитозоль за допомогою спеціальної транспортує системи.
У цитоплазмі цитрат реагує з HS-KoA і АТР, знову розпадаючись на ацетил КоА і оксалоацетат. Ця реакція каталізується АТР-цітратліазой.
Першою реакцією біосинтезу жирних кислот є карбоксилирование аце- тил-КоА, для чого потрібні бікарбонат, АТР, іони марганцю. Каталізує цю реакцію фермент ацетил-КоА-карбоксилаза. Фермент містить в якості про- стетичне групи біотин. Авидин - інгібітор біотину - пригнічує цю реакцію, як і синтез в цілому. Реакція протікає в два етапи: карбоксилирование біотину за участю АТР і перенесення карбоксильної групи на ацетил-КоА, в результаті чого утворюється Малоні-КоА:
Малоні-КоА є перший специфічний продукт біосинтезу жирних кислот. У присутності відповідної ферментної системи він швидко перетворюється в жирні кислоти.
Ферментні системи, що здійснюють синтез жирних кислот, називаються жирнокислотного синтетазами і діляться на 2 групи.
До першої групи належать поліферментні синтетази (тварини тканини і дріжджі). Друга група включає власне жирнокислотного сінтетези. Вони зустрічаються у ряду мікроорганізмів і рослин.
Поліферментний комплекс, який бере участь в синтезі вищих жирних кислот, складається з 6 ферментів, пов'язаних з так званим ацілпереносящім білком (АПБ). Цей білок термостабилен, має дві вільні HS-групи і втягується в процес синтезу вищих жирних кислот практично на всіх етапах. Молярна маса АПБ близько 10 000 г / моль. Даний білок в синтетазної системі виконує роль Ко А.
Послідовність реакцій, що відбуваються при синтезі жирних кислот, складається з 6 стадій:

Далі цикл реакцій повторюється. Завершується синтез жирної кислоти отщеплением HS-АПБ від ацил-АПБ під впливом ферменту діацілази.
Наприклад, в разі пальмітинової кислоти:
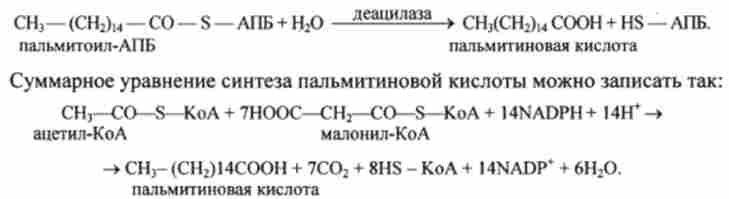
З огляду на, що на освіту однієї молекули Малоні-КоА і ацетил-КоА витрачаються 1 молекула АТР і одна молекула СО2, яка потім відщеплюється, сумарне рівняння можна представити і в такому вигляді:

Схема синтезу жирних кислот в кишкової палички Е. coli представлена на рис. 9.17.
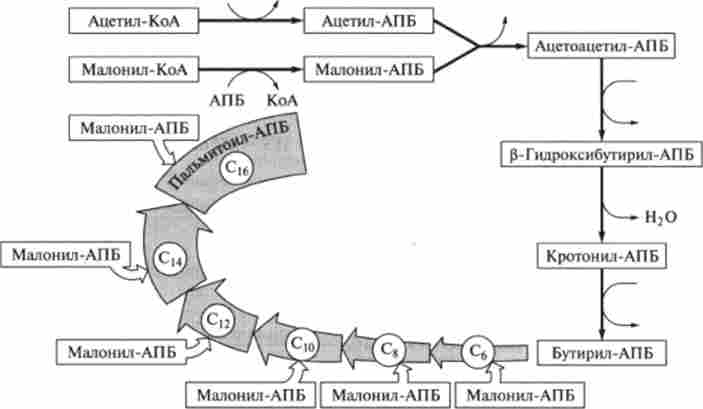
Мал. 9.17. Синтез пальмітинової кислоти в кишкової палички Е. coli (АНЬ - ацілпсрекосящій
білок)
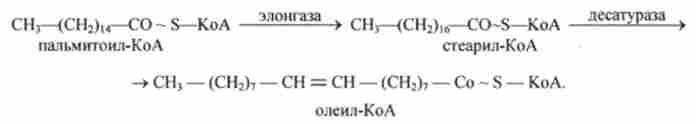
Тканини тварин, на відміну від рослинних тканин, мають досить обмежену здатність перетворювати насичені жирні кислоти в ненасичені. Це перетворення називається десатурація (освіта подвійних зв'язків).
Найбільш поширені мононенасичені жирні кислоти - пальміто- оленів і оленів - синтезуються з пальмітинової і стеаринової кислот. Цей синтез протікає в мікросомах клітин печінки і жирової тканини за участю молекулярного кисню. Перетворенню піддаються тільки активовані форми пальмітинової і стеаринової кислот. Ферменти, що беруть участь у перетворенні насичених жирні кислот в ненасичені, називають Десата разам і.
Поряд з десатурація жирних кислот в мікросомах відбувається і подовження їх ланцюгів (елонгація), причому обидва ці процеси можуть поєднуватися і повторюватися. Подовження ланцюга жирної кислоти відбувається шляхом послідовного приєднання 2-вуглецевих фрагментів до відповідного ацил-КоА за участю ма лоніл-КоА і NADPH. Ензиматична система, що каталізує подовження жирних кислот, отримала назву елонгази.
Наприклад, перетворення пальмітинової кислоти в реакціях елонгації і десатурации відбуваються наступним чином: