 гліцерину при дії гліцерин кінази:
гліцерину при дії гліцерин кінази:| Головна |
| «« | ЗМІСТ | »» |
|---|
В даний час показано, що в мікросомах клітин ссавців освіту подвійних зв'язків може відбуватися тільки на ділянці ланцюга жирної кислоти від 9-го до 1-го вуглецевого атома, бо в мікросомах відсутні десатурази, які могли б каталізувати утворення подвійних зв'язків в ланцюзі далі 9-го вуглецевого атома.
У тварин і рослин подвійні зв'язки можуть утворитися лише в певних положеннях вуглецевого ланцюга. Тому в організмі ссавців, в тому числі і людини, не можуть утворитися, наприклад, з стеаринової кислоти ліно- ліва і ліноленова кислоти, які відносяться до категорії незамінних жирних кислот.
До незамінних жирних кислот зазвичай зараховують також арахідонову кислоту '. У більшості ссавців вона може синтезуватися з л тюлевою кислоти.
Триацилгліцеридів (тригліцериди) грають в організмі роль запасних ліпідів. Вони активно утворюються в печінці і в жировій тканині, а також в стінці кишечника, де активність ферменту гліцеролкінази висока. Тригліцериди синтезуються з гліцерину і жирних кислот (в основному стеаринової, пальмітинової і олеїнової).
Гліцерофосфатного шлях біосинтезу тригліцеридів в тканинах проходить через освіту гліцерофосфату (гліцерин-3-фосфату) і коа-похідного жирної кислоти як проміжних сполук. Гліцерофосфат утворюється з двох різних джерел.
У нирках перша стадія синтезу тригліцеридів складається в ацилированием вільних гідроксильних груп глицерофосфата двома молекулами КоА- похідного жирної кислоти з утворенням фосфатидного кислоти.
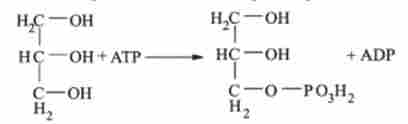
Гліцерофосфат може утворитися з
гліцерину при дії гліцерин кінази:
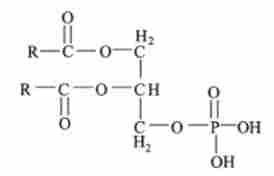
Потім при послідовному перенесення двох ацильних залишків виходить фосфатидними кислота:
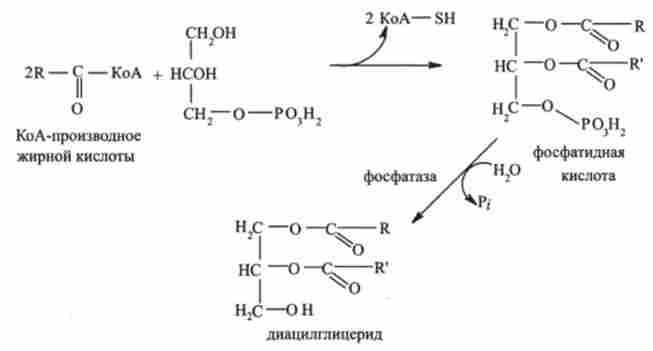
Далі від фосфатидного кислоти гидролитично отщепляется фосфорна кислота з утворенням діацілгліцерідов. У цій реакції беруть участь переважно насичені і ненасичені Cj6- і С ^ -проізводние КоА:
До утворився діацілгліцеріду приєднується залишок вищої жирної кислоти:
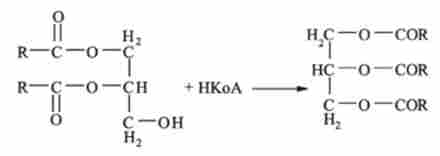
В результаті утворюється триацилгліцеридів (тригліцериди).
Перебіг того чи іншого шляху ресинтезу тригліцеридів залежить від складу продуктів розщеплення харчових ліпідів, що надійшли в кишкову стінку.
Клітини кишкового епітелію людини та вищих тварин активно синтезують тригліцериди в процесі всмоктування жирних кислот в тонкому кишечнику. Тут має місце інший тип реакції ацилювання. Моноацілгліцеріди, що утворюються в кишечнику вже при перетравленні харчових жирів, можуть аціліро- тися безпосередньо, минаючи стадію фосфатидного кислоти.
Гліцерофосфатного шлях набуває великого значення, коли в стінку кишечника надійшли переважно одні жирні кислоти. Якщо жирні кислоти надійшли в стінку з моногліцерид, запускається моногліцерідний шлях. При цьому надлишок моноглицеридов в епітеліальних клітинах гальмує протікання гліцсрофосфатного шляху.
Епітеліальні клітини кишечника здатні синтезувати фосфоліпіди і з вступників з їжею вільних жирних кислот, гліцерину і аміноспіртов. Цей процес можна розбити на три етапи:
В жировій тканині і м'язах внаслідок дуже низької активності гліцеролкі- називаються освіту гліцерин-3-фосфату в основному пов'язано з процесами гліколізу і глікогенолізу. Звичайним попередником гліцеролфосфата тут служить ди- оксіацетонфосфат, що утворюється в процесі гліколізу в результаті реакції, яка каталізується цитоплазматичної NAD-залежної гліцерофосфатдегід- рогеназой:
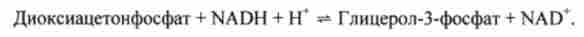
Відзначено, що якщо вміст глюкози в жировій тканині знижений (наприклад, при голодуванні), то утворюється лише незначна кількість гліцерин-3 фосфату і звільнилися в ході ліполізу вільні жирні кислоти не можуть бути використані для біосинтезу тригліцеридів. Тому жирні кислоти йдуть з жирової тканини. Навпаки, активація гліколізу в жировій тканині сприяє накопиченню в ній тригліцеридів, а також входять до їх складу жирних кислот. У печінки спостерігаються обидва шляхи утворення гліцерин-3-фосфату.
Встановлено, що більшість ферментів, що беруть участь в біосинтезі тригліцеридів, знаходяться в ЕПР і тільки деякі, наприклад гліцерин-3-фосфат-ацілтрансферази, в мітохондріях.