Синтез жирів в організмі відбувається головним чином з вуглеводів, що надходять в надмірній кількості і не використовуваних для синтезу глікогену. Крім цього, в синтезі ліпідів беруть участь також і деякі амінокислоти. У порівнянні з глікогеном жири більш компактну форму зберігання енергії, оскільки вони менш окислені і гідратованих. При цьому кількість енергії, резервувати у вигляді нейтральних ліпідів в жирових клітинах, нічим не обмежується на відміну від глікогену. Центральним процесом в Ліпогенез є синтез жирних кислот, оскільки вони входять до складу практично всіх груп ліпідів. Крім цього, слід пам'ятати, що основним джерелом енергії в жирах, здатним трансформуватися в хімічну енергію молекул АТФ, є процеси окислювальних перетворень саме жирних кислот.
Біосинтез жирних кислот
Структурним попередником для синтезу жирних кислот є ацетил-КоА. Це з'єднання утворюється в матриксі мітохондрій переважно з пірувату, в результаті реакції його окисного декарбоксілі- вання, а також в процесі р-окіслснія жирних кислот. Отже, вуглеводневі ланцюги збираються в ході послідовного приєднання двухуглсродних фрагментів в формі ацетил-КоА, т. Е. Біосинтез жирних кислот відбувається за тією ж схемою, але в протилежному напрямку порівняно з р-окіслсніем.
Однак існує ряд особливостей, що розрізняють ці два процеси, завдяки яким вони стають термодинамічно вигідними, незворотними і по-різному регулюються.
Слід зазначити основні відмінні риси анаболізму жирних кислот.
Синтез насичених кислот з довжиною вуглеводневого ланцюга до С16 (Пальмітинова кислота) в клітині здійснюється в цитоплазмі клітини. Подальше нарощування ланцюга відбувається в мітохондріях і частково в ЕПР, де йде перетворення насичених кислот в ненасичені.
Термодинамічно важливим є карбоксилирование ацетил-КоА і перетворення його в Малоні-КоА (СООН-СН2-СООН), на утворення якого витрачається одна макроергічних зв'язків молекули АТФ. З восьми молекул ацетил-КоА, необхідних для синтезу пальмітинової кислоти, тільки одна включається в реакції у вигляді ацетил-КоА, інші сім у вигляді Малоні-КоА.
В якості донора відновлювальних еквівалентів для відновлення кетогрупи до гідроксигрупи функціонує НАДФН, в той час як при зворотної реакції в процесі р-окислення відновлюється НАДН або ФАДН2в реакціях дегідрування ацил-КоА.
Ферменти, що каталізують анаболизм жирних кислот, об'єднані в єдиний Мультиферментний комплекс, який отримав назву «синтетаза вищих жирних кислот».
На всіх етапах синтезу жирних кислот активовані ацильні залишки пов'язані з ацілпереносящім білком, а не з коензимом А, як в процесі р-окислення жирних кислот.
Транспорт внутрімітохондріального ацетил-КоА в цитоплазму. Ацетил-КоА утворюється в клітці переважно в процесі всередині мітохондріальних реакцій окислення. Як відомо, мітохондріальна мембрана непроникна для ацетил-КоА.
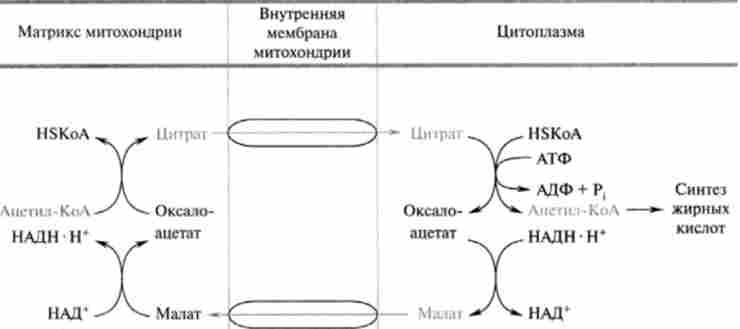
Відомі дві транспортні системи, що забезпечують перенесення ацетил-КоА з мітохондрій в цитоплазму: ацил-карнітіновий механізм, описаний раніше, і цитрат-транспортна система (рис. 23.14).
Мал. 23.14. Цитратний механізм транспорту ацетил-КоА через внутрішню мембрану мітохондрій
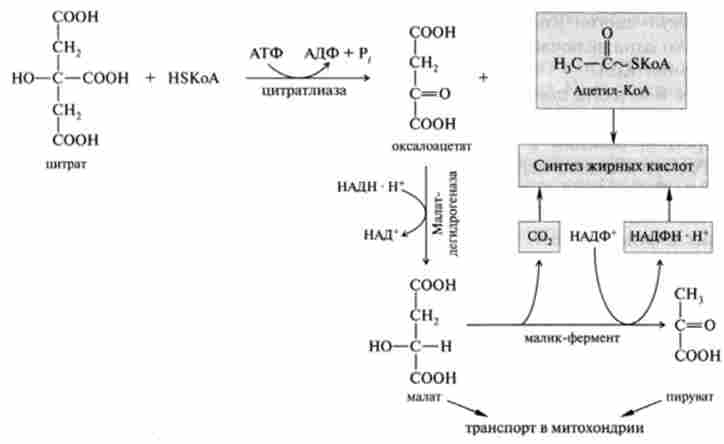
У процесі транспорту всередині мітохондріального ацетил-КоА в цитоплазму по нитратному механізму спочатку відбувається його взаємодія з оксалоацетата, який перетворюється в цитрат (перша реакція циклу три- карбонових кислот, що каталізується ферментом цітратсінтазой; гл. 19). Специфічною транслоказ утворився цитрат переноситься в цитоплазму, де розщеплюється ферментом цітратліазой за участю коензиму А на окса- лоацстат і ацетил-КоА. Механізм цієї реакції, пов'язаної з гідролізом АТФ, наведено нижче:
У зв'язку з тим що для оксалоацетата мембрана мітохондрії непроникна, вже в цитоплазмі він відновлюється за допомогою НАДН в малат, який за участю специфічної транслокази може повернутися в матрикс мітохондрії, де окислюється до оксалатацетата. Таким чином, завершується так званий човниковий механізм транспорту ацетил через метохондріальную мембрану. Частина цитоплазматичного малата піддається окислювальному дскарбоксілірованію і перетворюється в піруват за допомогою особливого «малик» - ферменту, коферментом якого є НАДФ+. Відновлений НАДФН поряд з ацетил-КоА і С02 використовується в синтезі жирних кислот.
! Зверніть увагу, що цитрат транспортується в цитоплазму лише тоді, коли його концентрація в матриксі мітохондрії досить велика, наприклад при надлишку вуглеводів, коли цикл трикарбонових кислот забезпечений ацетил-КоА.
Таким чином, цитратний механізм забезпечує як транспорт аце- тил-КоА з мітохондрії, так і приблизно на 50% потреби в НАДФН, який використовується в відновних реакціях синтезу жирних кислот. Крім цього, потреби в НАДФН заповнюються також за рахунок пентозофос- фатного шляху окислення глюкози.
Будову і функції клітини - вікова анатомія і фізіологія Клітини організму людини розрізняються за розмірами, формою, будовою і функціями (рис. 1.2), проте в їх структурі багато спільного. Так, кожна клітина має оболонку (плазматичну мембрану) і містить ядро і цитоплазму (рис. 1.3). У цитоплазмі знаходяться численні органели - лізосоми, мітохондрії,
Будову і функції жіночих статевих залоз - фізіологія людини і тварин Жіночі статеві органи представлені яєчниками - парними органами, розташованими в області малого тазу (рис. 3.4). За розмірами вони приблизно в два рази менше насінників, але, на відміну від них, не змінюють свого положення в протягом життя жінки. Мал. 3.4. Основні елементи будови зовнішніх
Будову біологічних мембран, хімічний склад - біохімія Мембрани прокаріотів і еукаріотів, так само як і мембрани тваринних і рослинних клітин, відрізняються один від одного по набору органел, складом і властивостями. Найбільш складноорганізованим є клітини еукаріот. У еукаріот виділяють наступні основні групи мембран: плазматичну, ядерну, ЕПР,
Будова хромосом. Каріотип - генетика в 2 Ч. Частина 1 Морфологію хромосом зазвичай описують на стадії метафази або анафази, коли вони краще видно в клітці. Для деяких рослин морфологію хромосом можна описати в профазі мітозу або мейозу. Залежно від розташування центромери розрізняють: а) акроцентрічні, або палочкообразниє, хромосоми, у яких центромера
Біосинтез замінних амінокислот - біохімія частина 2. Людина і тварини здатні синтезувати тільки 10 з 20 амінокислот, необхідних для синтезу білка, - це замінні амінокислоти (24.2). Шляхи біосинтезу цих амінокислот різноманітні, але при цьому вони володіють однією важливою властивістю: (Т) вуглецевий скелет амінокислот утворюється з проміжних