| Головна |
| «« | ЗМІСТ | »» |
|---|
Перший доказ ролі ДНК як носія спадкової інформації організмів привернув величезну увагу до вивчення нуклеїнових кислот. У 1869 р Ф. Мішер виділив з ядер клітин особливу речовину, яку назвав нуклєїнах. Через 20 років ця назва була замінена терміном нуклеїнова кислота. У 1924 р Р. Фельгена розробив метод цитологічного розпізнавання нуклеїнових кислот за допомогою їх специфічного фарбування і показав, що ДНК локалізується в ядрах клітин, а РНК - в цитоплазмі. У 1936 р А. Н. Білозерський і І. І. Дубровська виділили ДНК в чистому вигляді з ядер рослинних клітин. На початок 1930-х рр. були з'ясовані основні хімічні принципи будови цукрів нуклеїнових кислот, а в 1953 р створена структурна модель ДНК.
Основна структурна одиниця нуклеїнових кислот - нуклеотид, який складається з трьох хімічно різних частин, з'єднаних ковалентними зв'язками (рис. 5.2).
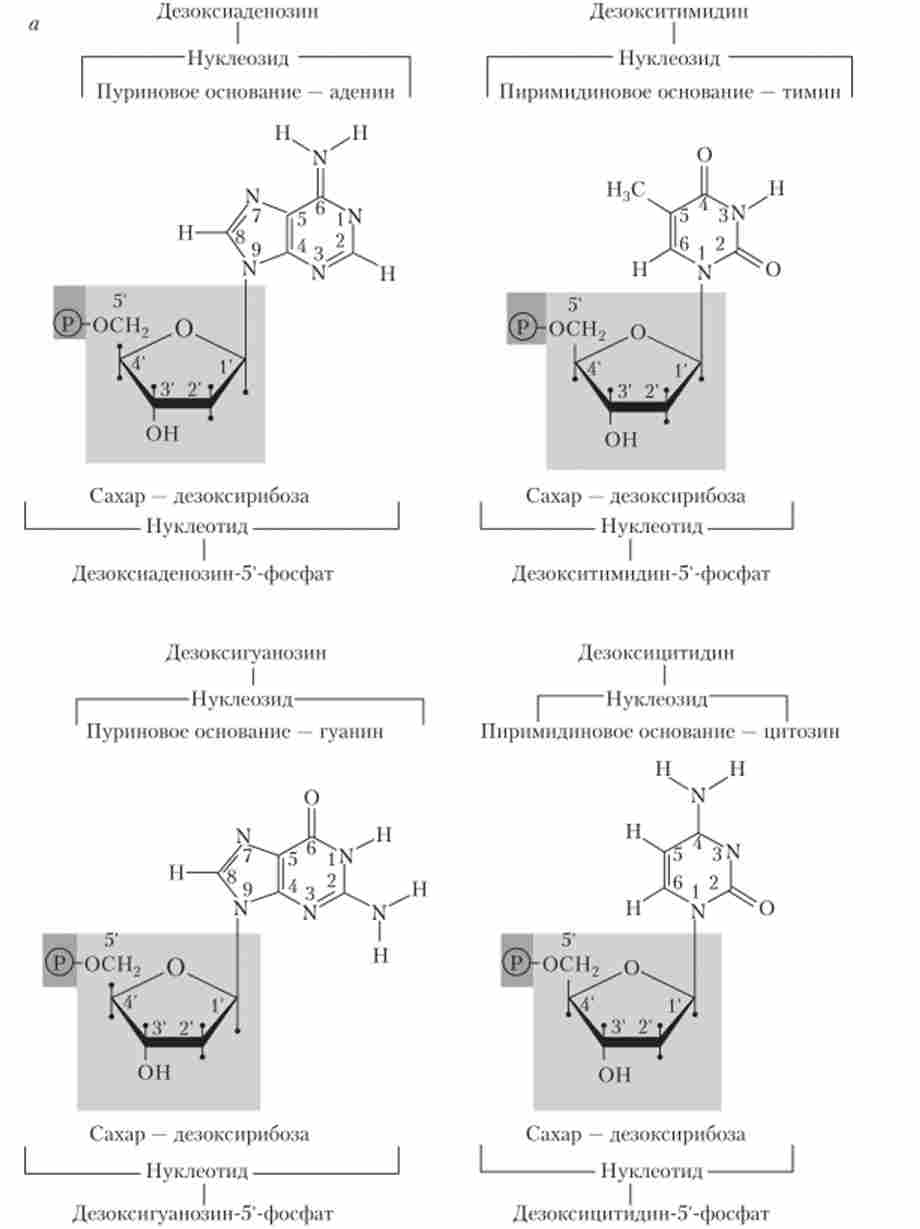
Мал. 5.2. Структурні формули: а - нуклеотидів; б - ДНК; в - РНК (див. Також с. 110)
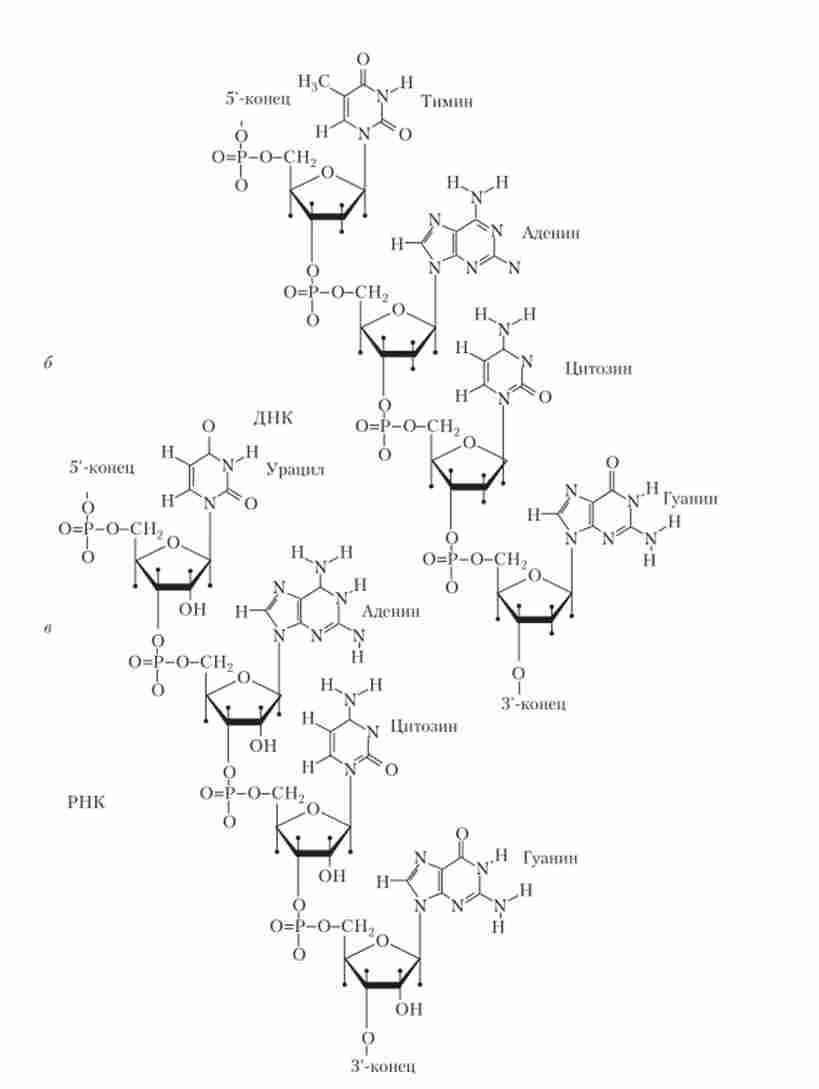
Мал. 5.2. Закінчення. Структурні формули: а - нуклеотидів; 6 - ДНК; в - РНК
Перша частина - цукор, що містить п'ять атомів вуглецю: дезоксірі- боза в ДНК і рибоза в РНК.
Друга частина нуклеотиду - пуриновое або пиримидиновое азотистих основ, ковалентно поєднане з першим атомом вуглецю цукру, формує структуру, яка називається нуклеозидом. ДНК містить пуринові основи - аденін (А) і гуанін (Г) - і піримідинові підстави - тимін (Т) і цитозин (Ц). Відповідні нуклеозиди називаються дезоксіаде- нозіном, дезоксигуанозину, дезокситимидина і дезоксіцітідіном. РНК містить ті ж пуринові основи, що і ДНК, пиримидиновое підставу цитозин, і замість тиміну в її склад входить урацил (У); відповідні нуклеозиди називаються аденозином, гуанозин, уридин і ци- тідіном.
Третю частину нуклеотиду становить фосфатна група, яка з'єднує сусідні нуклеозиди в полімерну ланцюжок за допомогою фосфо діефірних зв'язків між 5-атомом вуглецю одного цукру і З'-атомом вуглецю іншого (рис. 5.2, б, в). нуклеотидами називаються нуклеозиди з однією або декількома фосфатними групами, приєднаними ефірними зв'язками до 3'або 5-атомам вуглецю цукру. Синтез нуклеотидів передує синтезу нуклеїнових кислот, відповідно, нуклеотиди є продуктами хімічного або ферментативного гідролізу нуклеїнових кислот.
Нуклеїнові кислоти - дуже довгі полімерні ланцюжки, що складаються з мононуклеотидів, з'єднаних 5- і З'-фосфодіефірнимі зв'язками. Интактная молекула ДНК містить в залежності від виду організмів від декількох тисяч до багатьох мільйонів нуклеотидів, интактная молекула РНК - від 100 до 100 тис. І більше нуклеотидів.
Результати проведених Е. Чаргафф аналізів нуклеотидного складу ДНК різних видових форм показали, що молекулярне співвідношення різних азотистих основ - аденіну, гуаніну, тиміну, цитозину - варіює в широких межах. Отже, було доведено, що ДНК зовсім не монотонний полімер, що складається з однакових тетрануклеотідов, як припускали в 40-е гт. XX ст., І що він в повній мірі володіє складністю, необхідної для збереження і передачі спадкової інформації в формі специфічної послідовності нуклеотидних основ.
Дослідження Е. Чаргаффа виявили також особливість, притаманну всім молекулам ДНК: молярное зміст аденіну дорівнює змісту тиміну, а молярне зміст гуаніну - змістом цитозину. Ці рівності називаються правилом еквівалентності Чаргаффа: [А] = [Т], [Г] = [Ц]; кількість пуринів дорівнює кількості пиримидинов. Залежно від видової приналежності змінюється лише відношення ([А] + [Т]) / ([Г] + [Ц]) (табл. 5.1).
Зміст азотистих основ і їх співвідношення в ДНК різних організмів
|
Склад підстав, |
ставлення |
асиметрія |
|||||
|
об'єкт |
моль /% |
підстав |
|||||
|
А |
Г |
т |
ц |
А / Т |
г / ц |
(А + Т) / (Г + Ц) |
|
|
тварини |
|||||||
|
Людина |
30,9 |
19,9 |
29,4 |
19,8 |
1,05 |
1,00 |
1,52 |
|
вівця |
29,3 |
21,4 |
28,3 |
21,0 |
1,03 |
1,02 |
1,36 |
|
курка |
28,8 |
20,5 |
29,2 |
21,5 |
1,02 |
0,95 |
1,38 |
|
черепаха |
29,7 |
22,0 |
27,9 |
21,3 |
1,05 |
1,03 |
1,31 |
|
лосось |
29,7 |
20,8 |
29,1 |
20,4 |
1,02 |
1,02 |
1,43 |
|
морський краб |
47,3 |
2,7 |
47,3 |
2,7 |
1,00 |
1,00 |
17,50 |
|
Морський їжак |
32,8 |
17,7 |
32,1 |
17,2 |
1,02 |
1,02 |
1,58 |
|
Саранча |
29,3 |
20,5 |
29,3 |
20,7 |
1,00 |
1,00 |
1,41 |
|
Рослини, гриби |
|||||||
|
зародок пшениці |
27,3 |
22,7 |
27,1 |
22,8 |
1,01 |
1,00 |
U9 |
|
дріжджі |
31,3 |
18,7 |
32,9 |
17,1 |
0,95 |
1,09 |
1,79 |
|
гриб Aspergillus niger |
25,0 |
25,1 |
24,9 |
25,0 |
1,00 |
1,00 |
1,00 |
|
бактерії |
|||||||
|
Escherichia coli |
24,7 |
26,0 |
23,6 |
25,7 |
1,04 |
1,01 |
0,93 |
|
Staphylococcus aureus |
30,8 |
21.0 |
29,2 |
19,0 |
1,05 |
1.11 |
1,50 |
|
Clostridium perfringens |
36,9 |
14,0 |
36,3 |
12,8 |
1,01 |
1,09 |
2,70 |
|
Brucela abortus |
21,0 |
29,0 |
21,1 |
28,9 |
1,00 |
1,00 |
0,72 |
|
Sarcina lutea |
13,4 |
37,1 |
12,4 |
37,1 |
1,08 |
1,00 |
0,35 |
|
бактеріофаги |
|||||||
|
T7 |
26,0 |
24,0 |
26,0 |
24,0 |
1,00 |
1,00 |
1,08 |
|
X |
21,3 |
28,6 |
22,9 |
27,2 |
0,92 |
1,05 |
0,79 |
|
ФГ 174 (вірусна форма) |
24,6 |
24,1 |
32,7 |
18,5 |
0,75 |
1,30 |
1,34 |
|
ФГ 174 (репликативная форма) |
26,3 |
22,3 |
26,4 |
22,3 |
1,00 |
1,00 |
1,18 |
Ставлення підстав отримало найменування коефіцієнта нуклеотидной (Видовий) специфічності. У відкритті Чаргаффа була сформульована важлива структурна особливість ДНК, що знайшла пізніше відображення в структурній моделі ДНК Дж. Уотсона і Ф. Кріка (1953), які фактично показали, що правила Чаргаффа не накладаються ніяких обмежень на можливе число поєднань різних послідовностей підстав, здатних утворювати молекули ДНК.
Положення про нуклеотидної специфічності лягло в основу нової галузі біології - геносистематики, яка оперує порівнянням складу і структури нуклеїнових кислот для побудови природної системи організмів.
Відповідно до моделі Уотсона-Кріка молекула ДНК складається з двох по- лінуклеотідних ланцюжків (ниток, тяжів), з'єднаних один з одним за допомогою поперечних водневих зв'язків між азотистими підставами по комплементарному принципом (аденін одного ланцюга з'єднаний двома водневими зв'язками з тиміном протилежного ланцюжка, а гуанін і цитозин різних ланцюжків сполучені один з одним трьома водневими зв'язками). При цьому дві полінуклеотидних ланцюжка однієї молекули є анти- паралельними, т. Е. Навпаки 3'-кінця одного ланцюжка знаходиться 5'-кінець інший ланцюжка і навпаки (рис. 5.3). Слід, однак, мати на увазі сучасні дані про те, що генетичний матеріал деяких вірусів представлений одноланцюжковий (однониткових) молекулами ДНК. На підставі даних рентгеноструктурного аналізу ДНК Дж. Уотсон і Ф. Крик зробили також висновок про те, що її дволанцюжкова молекула має вторинну структуру у формі спіралі, закрученої в напрямку зліва направо, яка в подальшому отримала назву 5-форми (рис. 5.4). До теперішнього часу доведено, що крім найбільш часто зустрічається 5-форми можна виявити ділянки ДНК, які мають іншу конфігурацію, - як правозакрученной (форми А, С), так і закручену справа наліво (левозакрученной, або Z-форму) (рис. 5.4). Між цими формами вторинної структури ДНК є певні відмінності (табл. 5.2). Так, наприклад, відстань між двома сусідніми парами азотистих основ в двухцепочечной спіралі, виражене в нанометрах (нм), для 5-форми і Z-форми характеризується різними величинами (0,34 і 0,38 нм відповідно). На рис. 5.5 наведені сучасні об'ємні моделі «левозакрученной» і «правозакрученной» форм ДНК.
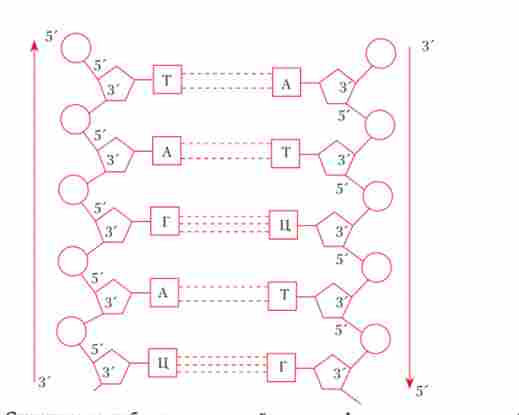
Мал. 5.3. схематичне ізоораженіе первинної структури фрагмента двухцепочечной молекули ДНК: А - аденін; Г - гуанін; Т - тимін; Ц - цитозин
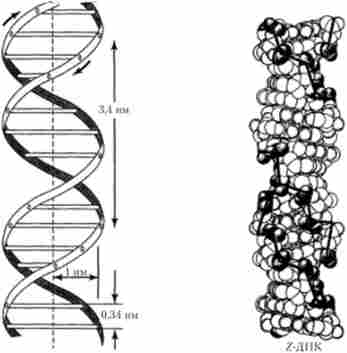
Мал. 5.4. Модель вторинної структури ДНК Рис. 5.5. Просторові моделі Уотсона-Кріка Z-форми і 5-форми ДНК

Таблиця 5.2
Властивості різних форм подвійних спіралей ДНК
|
властивості |
форми спіралей |
|||
|
А |
В |
З |
Z |
|
|
напрямок скрученности |
направо |
направо |
направо |
ліворуч |
|
Відстань між сусідніми парами підстав (нм) |
0,23 |
0,34 |
0,30 |
0,38 |
|
Число пар основ в одному витку спіралі |
10,7 |
10,0 |
9,3 |
12,0 |
|
Діаметр спіралі (нм) |
2,3 |
2,0 |
1,9 |
1,8 |
|
Кут нахилу підстав до осі спіралі (градуси) |
+19 |
-1,2 |
-6 |
-9 |
Молекули РНК в залежності від їх структурно-функціональних особливостей поділяють на кілька типів: інформаційні (матричні) РНК (іРНК, або мРНК), Хвороби (рРНК), транспортні РНК (тРНК), малі ядерні РНК (мяРНК) і ін. На відміну від ДНК молекули РНК завжди є одноланцюжковий (однониткових). Однак вони можуть формувати більш складні (вторинні) конфігурації за рахунок комплементарного з'єднання окремих ділянок такого ланцюжка на основі взаємодії комплементарних азотистих основ (A-У і Г-Ц). Як приклад можна розглянути конфігурацію, що має форму «трилисника», для молекули феніл ал аніновой транспортної РНК (рис. 5.6).
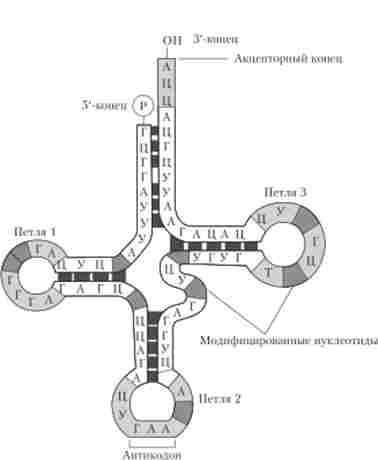
Мал. 5.6. Структура феніл ал аніновой тРНК дріжджів
У 1953 р Д. Уотсон і Ф. Крик запропонували модель структури ДНК, яка ґрунтувалася на наступних постулатах: