| Головна |
| «« | ЗМІСТ | »» |
|---|
Паралелізм в поведінці хромосом і генів наштовхнув дослідників на думку зіставити плоїдність (число наборів хромосом) клітин і зміст в них ДНК. В кінці 1940-х рр. А. Мирський і X. Рис показали, що в клітинах різних тканин одного і того ж організму кількість ДНК в розрахунку на гаплоїдний набір хромосом постійно, причому в статевих клітинах вона представлена в половинній кількості. Це можна вважати доказом ролі ДНК у спадковості.
Друге прямий доказ ролі ДНК у спадковості отримали в 1952 р А. Херші і М. Чейз при вивченні розмноження бактеріофага Т2 при інфікуванні кишкової палички Escherichia coli. Цей вірус складається з двох макромолекулярних компонентів: білка і ДНК. Остання укладена в білкову оболонку.
Частка бактеріофага схожа на пуголовка з булавовидний головкою і «хвостом» з відростками (рис. 5.7). При інфікуванні бактерії фаг Т2 приєднується за допомогою хвостових ниток до клітки Е. coli і «впорскує» в неї вміст своєї головки, т. е. молекулу ДНК без білка. Настає період розмноження частинок фага в клітині бактерії. Проникаючи в клітину бактерії, ДНК вірусу змінює функціонування генетичного апарату бактерії. ДНК фага кодує фермент, репресують гени бакгеріі. В результаті цього хромосома Е. coli руйнується, а ДНК-полімераза клітини бактерії використовується для реплікації вірусної ДНК. Інші компоненти фагової частки, синтезуючи окремо, виникають заново.
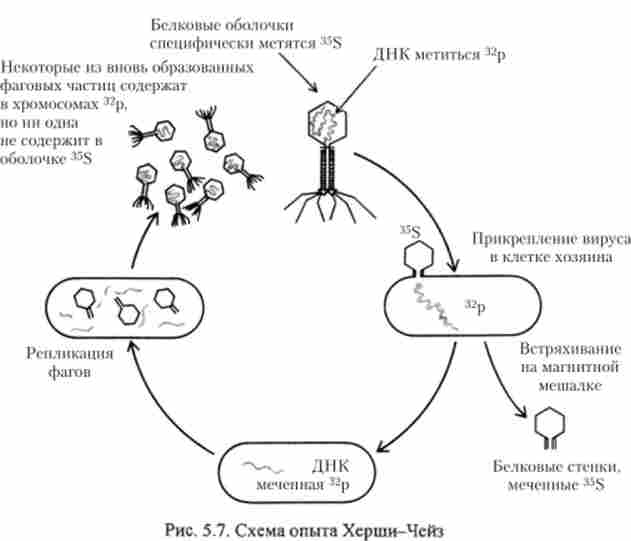
В експерименті А. Херші і М. Чейз була поставлена задача з'ясувати роль кожного з двох макромолекулярних компонентів (ДНК і білка) у формуванні потомства фага, використовуючи метод радіоактивних ізотопів. В якості маркера ДНК використовували радіоактивний фосфор, а білок мітили радіоактивної сіркою, що входить до складу амінокислот метіоніну і цистеїну. Фаг Т2 розмножували на бактеріях, що культивуються в середовищі з радіоактивним ізотопом 35S, в результаті чого білок фага був позначений цим ізотопом. Щонайменше 99% всього фосфору в фаге Т2 доводиться на частку ДНК; її позначили радіоактивним ізотопом 32Р. Ці радіоактивні мітки дозволяли простежити шляхи білка і ДНК фага Т2 при інфекції.
Інфекційний процес починається з прикріплення фага до бактеріальної клітці (рис. 5.7). Цей етап можна спостерігати за допомогою електронного мікроскопа.
Результати спостережень підтверджуються тим, що при центрифугуванні клітин на даній стадії інфекції фаги, що містять як 35S, так і 32Р, осідають разом з бактеріями. А. Херші і М. Чейз виявили, що незабаром після інфікування більшу частину міченого 35S білка можна відокремити від бактеріальних клітин, активно перемішуючи і струшуючи культуру на мішалці; проте велика частина меченной 32Р ДНК не відділяється при цьому від бактеріальних клітин, оскільки, по-видимому, виявляється в цей час вже всередині них. Усунення з культури порожніх білкових оболонок фага ( «тіней») не впливає на подальші події: відбувається лізис бактерій, і з них виходить потомство фага точно так же, як в тому випадку, коли тіні залишаються прикріпленими до клітин (рис. 5.7). На підставі результатів цього досвіду А. Херші і М. Чейз зробили висновок, що для утворення копій фага в зараженій бактеріальної клітці істотна лише ДНК батьківського фага, хоча самі копії містять як ДНК, так і білок. Авторами було висловлено припущення, що білковий компонент фага лише захищає ДНК від розщеплюють ферментів і забезпечує її потрапляння в бактеріальну клітину, тоді як ДНК являє собою власне речовина спадковості. Таким чином, доведено, що саме ДНК, а не білок визначає розмноження бактеріофага в інфікованих клітинах бактерії.