| Головна |
| «« | ЗМІСТ | »» |
|---|
Кишкова паличка {Escherichia coli) - бактерія, яка зазвичай мешкає в кишечнику людини і тварин. У сучасних дослідженнях з молекулярної генетики вона займає одне з перших місць. Це палочковидний організм близько 2 мкм завдовжки і 0,8-1 мкм завтовшки. Зовні клітина кишкової палички одягнена твердою оболонкою, зберігає свою форму навіть у тому випадку, якщо в силу тих чи інших причин втрачається її вміст. Під клітинною оболонкою лежить еластична тонка цитоплазматична мембрана. Якщо видалити клітинну оболонку дією на неї ферменту, то клітина разом зі своїм вмістом приймає форму кульки, званого протопластом, або сферопласти. Вміст клітини бакгеріі представлено протоплазми, що складається з цитоплазми і включених в неї молекул ДНК. Клітка гаплоидное і містить одну хромосому, що представляє собою довгу кільцеву нитка ДНК. Її довжина близько 1 300 мкм, т. Е. Вона більш ніж в 600 разів довше самої бактеріальної клітини. Однак в клітці ДНК багаторазово згорнута і щільно «упакована». Щонайменше в одній точці хромосома фіксована на утвореннях, що відходять від цитоплазматичної мембрани в глиб клітки і званих Мезосоми.
Хромосома у кишкової палички реплицируется від однієї строго певної точки, в якій вона прикріплена до цитоплазматичної мембрани. «Хвилі реплікації» йдуть в обох напрямках від стартової точки і зустрічаються один з одним посередині хромосомного кільця. На реплікацію хромосоми у кишкової палички витрачається близько 20 хв. Потім синтезовані хромосоми відходять один від одного, і між ними починає формуватися перегородка з цитоплазматичної мембрани. Деякий час розмежовані перегородкою протопластів залишаються під одним клітинної оболонкою, потім клітина ділиться.
Дослідження показали, що у бактерії існує і статевий процес, що веде до об'єднання і подальшої рекомбінації генів різних батьківських типів. В основі статевого процесу у бактерій лежить кон'югація - поява тимчасової зв'язку між клітинами бактерій за допомогою освіти цитоплазматичного містка. Такий зв'язок створює умови для проникнення генетичного матеріалу з однієї клітини бактерії в іншу.
У 1952 р В. Хейс виявив, що при явищах рекомбінації у бактерій одна з ліній служить донором, а інша - реципієнтом. Донорні клітини (чоловічі клітини бактерій) характеризуються наявністю у них особливого фактора F * - не пов'язаної з хромосомою ДНК, здатної до самовідтворення. При кон'югації бактерій фактор F може переходити в реці- піентную жіночу клітку F ~, перетворюючи її в чоловічу.
Статевий фактор бактерій (F4} Містить 105 пар нуклеотидів, що відповідає за кількістю одному геному фага. Бактерії т. Е. Позбавлені статевого фактора, є реципієнтами. При кон'югації клітин F4 і F ~ в останню часто переходить тільки статевий фактор. Однак, коли фактор F4 в клітці донора з'єднується з се хромосомою, кінець останньої проникає через цитоплазматичний місток в клітку реципієнта, обумовлюючи появу клітин, здатних до особливо високій частоті рекомбінації. Ці клітини були позначені символом Hfr (Від англ. High frequency of recombination - висока частота рекомбінації).
На прикладі фактора F у бактерій було показано, що окрема ДНК може як існувати самостійно, так і бути включеною в хромосому бактерії у вигляді її частини. Такі молекули ДНК отримали назву Епіс.
Спочатку наявність кон'югації клітин бактерій постулировалось на основі генетичних експериментів, в яких демонструвалося явище рекомбінації, потім факт кон'югації бактерій був підтверджений за допомогою електронного мікроскопа. На рис. 5.8 ясно видно цитоплазматичний місток, який служить для переходу фактора F * і хромосоми бактерій з однієї клітини в іншу при їх кон'югації.
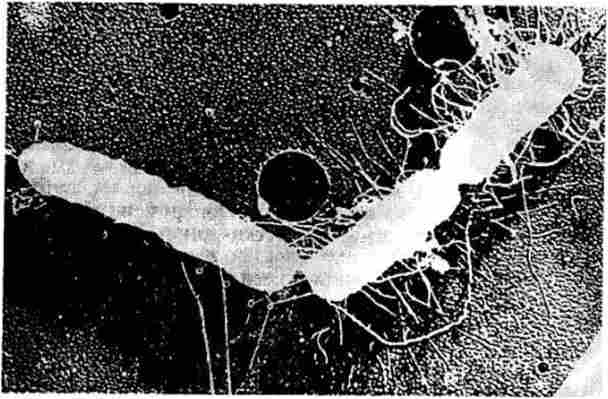
Мал. 5.8. Електронна мікрофотографія кон'югації між бактеріальними клітинами Hfr і F ~