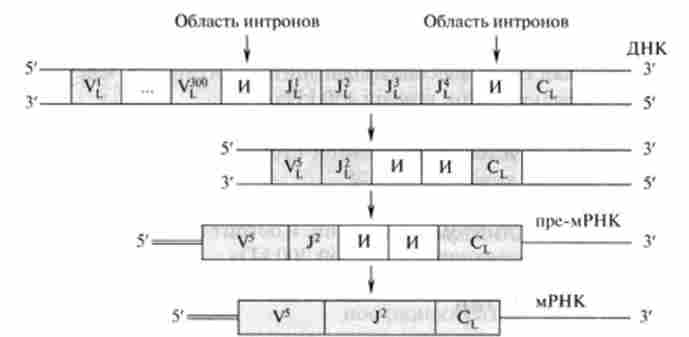
Величезна різноманітність антитіл неадекватно числу генів, локалізованих в лімфоцитах. Наприклад, в клітинах людини міститься не більше 105 генів, а число вироблюваних антитіл на 1-2 порядки більше. Імуноглобуліни є білками, отже, вони кодуються відповідними генами. Таким чином, розгадку феномену цієї невідповідності слід шукати в особливостях функціонування генома лімфоцитів. оказ&юсь, що синтез антитіл кодується трьома різними родинами незчеплених генів, розташованих в різних хромосомах. Розглянемо, як формується функціонально активна ділянка ДНК, відповідальний за утворення легкої ланцюга Ig. Виявлено близько 300 генів, що кодують варіабельні ділянки L-ланцюга (VL), один ген, КОЛІР синтез константного ділянки (CL), і від одного до чотирьох генів, відповідальних за синтез амінокислотних послідовностей, що з'єднують константні і варіабельні ділянки легкого ланцюга (J,) .
На рис. 30.10 представлена схема формування функціонально активних генів, що кодують легкий ланцюг Ig.
До початку перегрупування на матриці ДНК знаходиться до 300 генів, здатних кодувати варіабельні ділянки антитіл, а також до чотирьох генів з'єднання (J).
Перегрупування генів в даному випадку є спонтанний процес, в результаті чого кожен з 300 варіабельних генів випадково знаходить ген J і з'єднується з ним. Це з'єднання супроводжується вирізанням интронов на ланцюзі ДНК. Взаємодія 300 VL-reHOBс чотирма J-генами да-
Мал. 30.10. Освіта функціонально активних генів і мРНК, КОЛІР легкі ланцюги Ig ет 1200 різних комбінацій освіти активних генів. Це число значно збільшується за рахунок зрушень рамки вирізання генів, а також в результаті точкових мутацій в області варіабельних генів. Дані механізми характерні і для варіабельних ділянок важких ланцюгів. Утворилися комбінації антиген-связиваюшим центрів Ig мають різне спорідненість до антигену. Далі відбувається відбір, причому в якості селекціонера виступає антиген. В результаті утворюються плазматичні клітини, які синтезують імуноглобуліни з максимальним спорідненістю до даного антигену.
Будова серця - вікова анатомія і фізіологія. Т.2 опорно-рухова і вісцеральні системи 4 р діафрагма Мал. 9.2. Положення серця в грудній порожнині Серце розташоване в грудній порожнині між легкими і зазвичай зміщений вліво (рис. 9.2). Серце розташовується асиметрично: його вісь спрямована праворуч-зверху, вперед-вниз і вліво. Своїм підставою серце звернено до хребта, а сто верхівка
Будова нервової системи, спинний мозок - анатомія центральної нервової системи В результаті вивчення даного розділу студент повинен: знати морфологічний і клітинну будову СМ, основні області і зони (роги сірої речовини і канатики білої речовини); основні функції СМ, обумовлені зв'язками з областями ( «поверхами») тіла людини і з ГМ; основні види рефлекторних дуг СМ (моно-
Будова клітини - нервова система: анатомія, фізіологія, Нейрофармакологія Клітини різних організмів, що живуть на Землі, і навіть одного і того ж організму можуть дуже сильно відрізнятися і за розмірами, і але будовою, і але функціям. Наприклад, порівняйте нейрон, клітку серцевого м'яза, еритроцит і сперматозоїд (рис. 1.13). Мал . 1.13. Приклади клітин різної форми:
Будова дихальної системи - вікова фізіологія і психофізіологія Дихальна система складається з дихальних шляхів, респіраторного відділу легень, грудної клітини (включаючи її кістково-хрящової каркас і нервово-м'язову систему), судинної системи легенів і нервових центрів регуляції дихання (рис. 6.1). Мал. 6.1. Схема розташування органів дихальної системи
Блокада нервово-м'язової передачі - нейрофізіологія На прикладі роботи нервово-м'язового синапсу можна розглядати функціонування будь-якого хімічного синапсу. Також на цьому прикладі можна розглянути механізми придушення (гноблення) передачі сигналу в хімічному синапсі. Так, наприклад, існує кілька способів блокади нервово-м'язової передачі
Біосинтез нуклеотидів - біохімія частина 2. Майже всі організми здатні синтезувати піримідинові і пуринові нуклеотиди dc novo з простих сполук. Першим продуктом нуклеотидной природи пуринового шляху є і11озі11-5-монофосфат (ІМФ), що перетворюється потім в усі інші пуринові нуклеотиди. Структурним попередником всіх піримідинових нуклеотидів
Біосинтез дезоксирибонуклеотидів, протиракова терапія - біохімія частина 2. Для біосинтезу ДНК в якості субстратів необхідні дезоксірібонук- леотіди (дРНТ): дАТФ, ДГТФ, дЦТФ, дТТФ. Дезоксінуклсозідтріфосфати утворюються шляхом безпосереднього відновлення відповідних рибо нуклеозідполіфосфатов в ході процесу, для якого необхідні наступні умови: - субстрати для відновлення