| Головна |
| «« | ЗМІСТ | »» |
|---|
Для біосинтезу ДНК в якості субстратів необхідні дезоксірібонук- леотіди (дРНТ): дАТФ, ДГТФ, дЦТФ, дТТФ. Дезоксінуклсозідтріфосфати утворюються шляхом безпосереднього відновлення відповідних рибо нуклеозідполіфосфатов в ході процесу, для якого необхідні наступні умови:
- субстрати для відновлення - рібонуклеотіддіфосфати: АДФ, ГДФ, ЦДФ, УДФ;
* Безпосередній відновник рибонуклеотидов - білок тіоре- доксін;
два ферменти рибонуклеотидредуктазу (РНР) і тіоредоксінредуктази (ТРР);
наявність відновленого НАДФН - донора відновлювальних еквівалентів в реакції регенерації дисульфідній форми тіоредоксіна в сульфгід- р мулових.
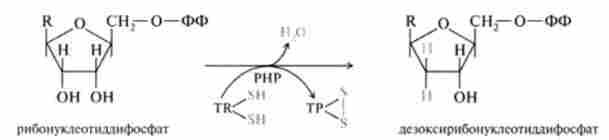
Відновлення рибонуклеотидов в дезоксирибонуклеотидів зводиться до елементарного акту - відновлення рибози в 2-дезоксирибози, що вимагає наявності двох атомів водню; безпосереднім джерелом відновлювальних еквівалентів виявився термостабільний білок - тіорелоксін. з-
sh
тримає дві вільні SH-групи (TRgp | J:
Тіорелоксін являє собою невеликий білок (11,7 kDa), що складається з однієї поліпептидного ланцюга з одного внутріцепочечной дисульфид ної
зв'язком (TRc ^, або TR-S2J. Фермент тіорсдоксінрсдуктаза (ТРР) каталізує НАДФН-залежне відновлення: -S-S - »- -SH + HS- з образо-
S н
ванием відновленого тіоредоксіна, або TR- (SH)2
Послідовність перетворень рібонуклеозіддіфосфатов в процесі синтезу дезоксірібонуклеозіддіфосфатов представлена на рис. 26.4.
Утворені в процесі біосинтезу дАДФ, дГДФ, дЦДФ фосфорілізу- ються в нуклеотідкіназной реакції до дАТФ. ДГТФ, дЦТФ; дУДФ є попередником дТТФ.
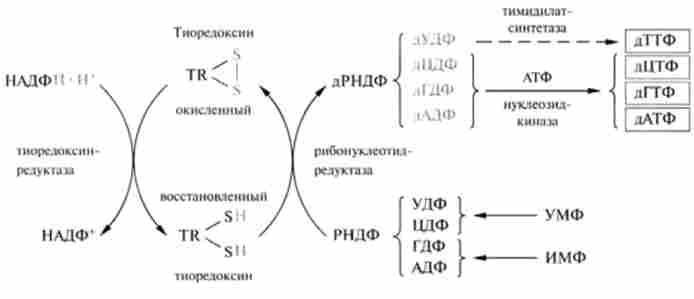
Мал. 26.4. Схема біосинтезу лезоксірібонуклеотідов:
РНДФ - рібонуклеозіддіфосфат; дРНДФ - дезоксірібонуклеозіддіфосфат
Біосинтез дТТФ. Цей процес здійснюється в три етапи. На першому етапі дУДФ гідролізується до дУМФ:
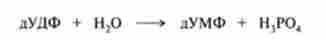
На другому етапі відбувається метилування дУМФ, де в якості донора метильної групи виступає метилентетрагідрофолат (N5-N,0-MeTH- льон-ТГФ), реакцію каталізує тіміділатсінтаза:
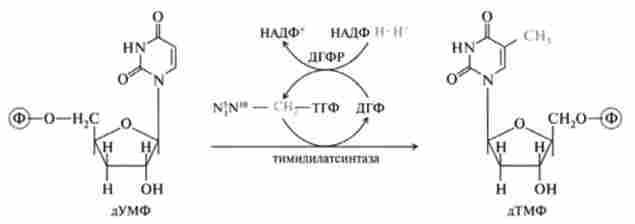
У цій реакції тетрагідрофолат перетворюється в дигідрофолат (ДГФ), так як N5- 1М10-метилен-ТГФ одночасно з метиленової групою (СН2) - віддає протон на її відновлення до СН3-групи. ДГФ регенерує до ТГФ під дією НДДФН-залежною дегідрофолатредуктази (ДГФР).
Третій етап включає дві стадії фосфорилювання, каталізуються ферментами нуклеозідмонофосфат- і нуклеозіддіфосфаттрансферамі за схемою:
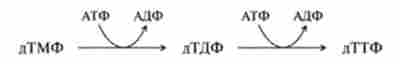
Недолік тиміну або тетрагідрофолату в швидкозростаючих і діляться пухлинних клітинах зупиняє клітинний ріст і навіть призводить до загибелі клітин. Інгібування дегідрофолатредуктази побічно пригнічує перетворення дУМФ -? дТМФ. На цьому заснована дія групи антіфолатних засобів, аналогів ТГФ, що мають протипухлинну дію. Особливо ефективні два препарати - аміноптерин і метотрексат.
Тіміділатсінтетазу - фермент, що каталізує реакцію дУМФ -? -? дТМФ, безпосередньо відзначено зниження 5-фтороурацілом і 5-фтор-2'-дез- оксіурідіном (конкурентне інгібування). Специфічне інгібування синтезу тиміну, необхідного для синтезу ДНК, також використовується в протиракової терапії.