| Головна |
| «« | ЗМІСТ | »» |
|---|
Для зорової системи характерним є досить складний дорецепторпий апарат, представлений оптичною системою ока (рис. 13.1) і різними допоміжними утвореннями. До них відносяться:
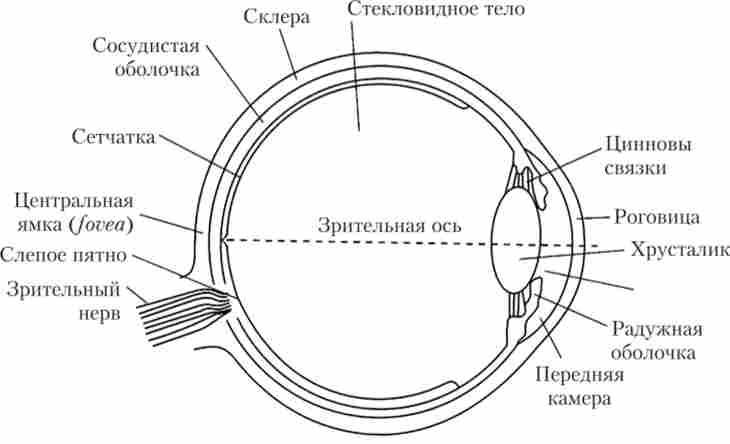
Мал. 13.1. будова очі1
Коротко зупинимося па основні функції елементів органу зору.
м'язів. При порушеннях процесу акомодації очі виникають такі захворювання, як міопія (Короткозорість) або гиперметропия (Далекозорість).
6. Скловидне тіло - також є частиною оптичної системи очі. Це колоїдний розчин гіалуронової кислоти (драглиста рідина).
В цілому оптична система ока забезпечує фокусування зображення на рецепторной поверхні сітківки. При цьому зображення потрапляє на сітківку дійсне (не спотворене), різко зменшене і перевернуте.
Самі рецептори розташовуються в сітківці (Рис. 13.2), яка має досить складну будову і є високоорганізовану шарувату структуру, що об'єднує не тільки рецептори, але і ряд інших нейронів. За складністю організації сітківку розглядають як частину мозку, винесену на периферію. У ній відбувається первинна обробка зорових сигналів, перетворення їх в нервові імпульси, що передаються в центральні структури.
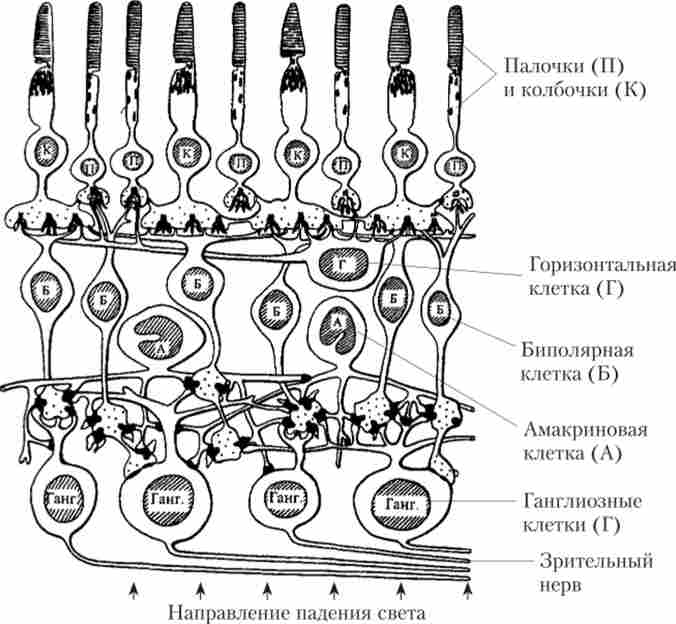
Мал. 13.2. Будова сітківки ока1
7. фоторецептори (Палички і колбочки) розташовані в сітківці, в місці найбільш віддаленому від кришталика, вони повернені від пучка падаючого світла.
палички відповідають за зір в темряві і сутінках (чорно-біле) за рахунок наявності в них пігменту родопсина. Їх в сітківці міститься приблизно 120 млн. колб в сітківці менше (приблизно 6 млн), і вони відповідальні за колірний зір завдяки наявності в них трьох типів зорових пігментів (йодопсин і ін.). зорові пігменти (Родопсин і йодопсин) складаються з ретиналю (альдегіду вітаміну Л) і гликопротеида онсіна. Вони близькі але будовою, але відрізняються за спектрами поглинання світлових хвиль: для родопсина, палочкового пігменту, максимум знаходиться на довжині хвилі близько 500 нм, а для йодопсіна, колбочкового пігменту, існує три піки в залежності від типу колбочки (430- 470 нм - синій колір, 500-530 їм - зелений, 620-760 їм - червоний). нестача вітаміну А в їжі призводить до порушення синтезу зорових пігментів і, як наслідок, до порушення сутінкового зору (куряча сліпота).
Механізм роботи зорового рецептора
Зовнішні сегменти фоторецепторів (паличок і колбочок) містять високочутливу багатоступеневу систему посилення сигналу в сітківці.
Внутрішньоклітинна реєстрація електричних процесів від фоторецепторів показала, що в темряві вздовж фоторецептора з внутрішнього до зовнішнього сегменту тече так званий темновой ток і безперервно йде виділення медіатора (рис. 13.3). Освітлення призводить до блокади цього струму. У темряві також відбувається ресіітез (відновлення) зорових пігментів, що розпалися під час висвітлення. Причому відновлення йодопсіна відбувається в 500 разів швидше, ніж родопсина. Цим пояснюються відмінності в швидкості світлової та темнової адаптації зорової системи.
Фоторецептори пов'язані між собою електричними синапсами (щілинними контактами), причому палички з паличками, а колбочки з колбами. Завдяки такому з'єднанню сигнал, що виник в одному рецепторі, швидко поширюється до сусідніх клітин.
В результаті складних фотохімічних процесів в фоторецепторах при дії світла (рис. 13.4) виникає рецепторний потенціал (РП) у вигляді гіпернолярізаціі мембрани рецептора. Така форма рецепторного потенціалу є винятком, так як у всіх інших рецепторних клітинах РП є деполяризацию мембрани сенсорної клітини. Однак, як і в випадку інших сенсорних систем, амплітуда гіперполярізаціонного РП зорових рецепторів зростає зі збільшенням інтенсивності освітлення.
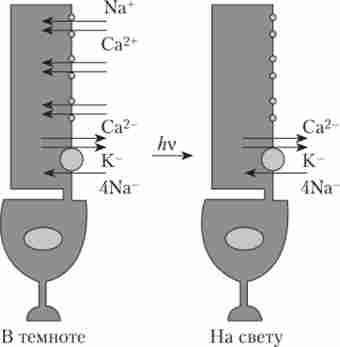
Мал. 13.3. Робота фоторецептора
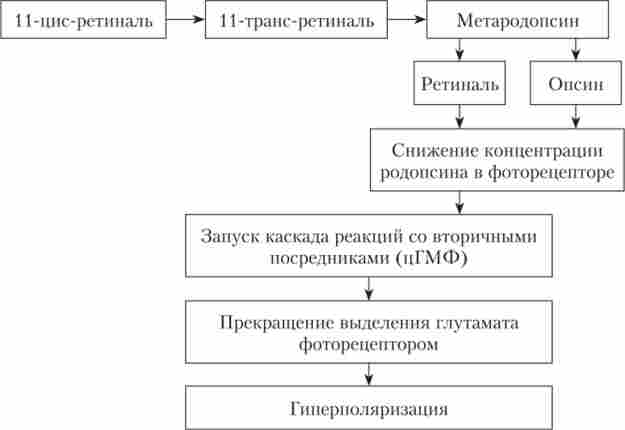
Мал. 13.4. Каскад реакцій при дії світла на фоторецептор
Отже, фоторецептори - це вторинні рецепторні клітини, їх відростки з'єднані з біполярними клітинами, а ті, в свою чергу, утворюють синапси з гангліозними клітинами. Аксони гангліозних клітин утворюють зоровий нерв.
Починаючи з рівня біполярних клітин, нейрони зорової системи формують концентричні рецептивні поля з центром і периферією, що знаходяться в антагоністичних відносинах.
Рецептивное иоле - це вся сукупність рецепторів, що посилають сигнали до одній клітці іншого рівня.
У сітківці такі рецептивні поля організовані таким чином, що якщо центр на освітлення реагує збудженням, то периферія гальмуванням (ои-центр і про // - периферія), або ж навпаки {off-центр і ои-периферія). Вважається, що цей механізм забезпечує можливість сприйняття двох протилежних класів зорових образів: світлі об'єкти на темпом фойє і темні об'єкти на світлому фойє.
10. гангліозних клітини є виходами з сітківки, саме їх довгі аксони формують зоровий нерв. Більшість гангліозних клітин мають концентричні рецептивні поля з центром і периферією по on- і р ^ -типу (рис. 13.5): при освітленні однієї зони ганглиозная клітина збуджується, а при її затемненні гальмується (ои-ефект), або ж навпаки (про ^ -ефект). Завдяки двом типам гангліозних клітин (з on- і райцентрами рецептивних полів) виявлення світлих і темних об'єктів в поле зору відбувається вже на рівні сітківки ока.
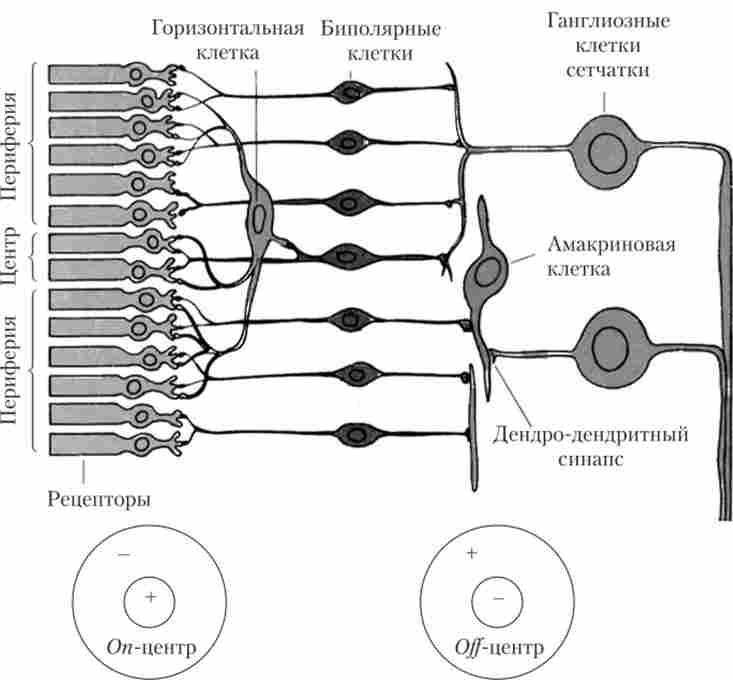
Мал. 135. Рецептивні поля гангліозних клітин сітківки з центром і периферією2
Ця схема (див. Рис. 13.5) показує, як могли б створюватися рецептивні поля з центром і периферією. Центр (в даному випадку про ^ -цснтр) утворює невелика група рецепторів, що мають потужні збуджуючі синапси з одиночної біполярної клітиною. Одна або кілька таких клітин з'єднані з гангліозних клітиною, формуючи центр її рецептивного поля. Периферію рецептивного поля біполярної клітини створює значно більшу кількість рецепторів, сполучених з горизонтальною кліткою збудливими синапсами.
У сітківці також існують два типи гальмівних нейронів: горизонтальні і амакріновие клітини.
11. Горизонтальні і амакріновие клітини. Горизонтальні клітини пов'язують фоторецептори з біполярними клітинами і можуть передавати сигнали уздовж зовнішнього синаптичного шару сітківки. Амакріновие клітини діють аналогічно горизонтальним, але тільки на рівні передачі сигналів від біполярних клітин до гангліозних клітинам. Горизонтальні і амакріновие клітини є гальмівними нейронами, вони забезпечують процеси латерального гальмування в сітківці.