| Головна |
| «« | ЗМІСТ | »» |
|---|
Плазміди є екстрахромосомние генетичні елементи (кільцеві молекули ДНК), які в бактеріальних клітинах фізично відокремлені від хромосоми і здатні довго підтримуватися і відтворюватися в такій формі. За розмірами дволанцюжкові молекули плазмідних ДНК близькі до генетичного матеріалу відповідних вірусів, і їх довжина зазвичай коливається від 0,5 до 100 мкм (молярна маса 1-200 МД).
Подібно до вірусів деякі плазміди можуть інтегруватися в бактеріальну хромосому за допомогою механізму сайт-специфічної рекомбінації і тривалий час існувати як складова частина її структури. При виході зі складу хромосоми під впливом тих чи інших індукують агентів, як і дтя вірусів, можливо правильне або неправильне «вирізання» плазміди. В іншому випадку в хромосомі залишаються фрагменти гшазмідного генома, а плазміда набуває ділянки ДНК (окремі гени) бактеріальної хромосоми. Разом з тим на відміну від вірусів плазміди не здатні руйнувати клітини, в яких вони знаходяться, т. Е. У них немає литичного варіанти розвитку.
Плазміди, що мають значні молекулярні розміри (понад 10 мкм завдовжки), виявляються в бактеріальних клітинах в невеликій кількості (1-3 копії на хромосому), тому їх називають малокопійни. ми плазмидами. З іншого боку, самостійна (автономна) реплікація дрібних плазмід (0,5-10 мкм) схильна до менш суворому контролю з боку хромосомної генетичної системи клітини, і їх число може досягати декількох десятків копій на хромосому, т. Е. Вони є многокопій- ними . Невеликі многокопійние плазміди являють собою зручний об'єкт для генетичної інженерії, і їх часто використовують в якості векторів при клонуванні генетичного матеріалу різних організмів.
Як і ДНК бактеріальної хромосоми, плазміди знаходяться в клітці в компактній суперспіралізованной формі, лінійні розміри якої значно менше первинних розмірів складової її плазмідної ДНК. Подібно хромосомної ДНК, при ферментативної обробці препарату плазмідної ДНК, виділеної з бактерій, можуть бути отримані відкриті кільцеві варіанти цих молекул, які дуже зручні для їх вивчення та визначення лінійних розмірів.
Генетична організація плазмід найбільш повно була досліджена на прикладі статевого фактора F (Фактора фертильності, фактора переносу), який виявився першою з ідентифікованих плазмід і був виявлений в клітинах штаму Е. coli К12. В даний час є відомості про локалізацію в складі плазміди F більше 60 різних генів, в тому числі відповідальних за її автономну реплікацію в клітці (генів гер), здатність забезпечувати перенесення генетичного матеріалу в процесі кон'югації бактерій (генів tra), Несумісність з іншими плазмідами (генів inc) та ін.
У структурі плазміди F, представляє собою кільцеву молекулу ДНК величиною 100 тис. пар нуклеотидів (100 т. п. н.), є три генетичні області реплікації (Rep FIA, Rep FIB, Rep FIC) (Рис. 5.36). Однак область Rep FIC інактивована вставкою транспозона (Тп 1000). У складі цієї плазміди постійно знаходяться також инсерционно послідовність IS2 і дві послідовності / S3.
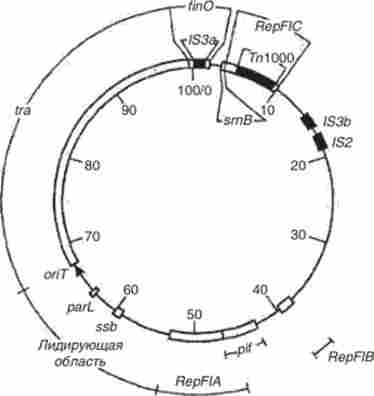
Мал. 5.36. Генетична карта плазміди F
Одне з основних властивостей плазміди F полягає в тому, що вона забезпечує містить її бактеріальним клітинам здатність бути донорами генетичного матеріалу (кон'югатівних), т. е. можливість вступати в кон'югацію з безплазмідних (реципієнтную) клітинами і передавати їм свою генетичну інформацію. Це властивість зазначеної плазміди і інших кон'югатівних (трансмісивних) плазмід детермінується знаходиться в їх складі генетичної областю tra (Від англ, transfer - перенос), що включає більше 20 rra-генів. У структурі плазміди F / Гд-область займає безперервний сегмент ДНК протяжністю 33,3 т. П. Н., Який починається в точці 66,7 т. П. Н. і закінчується в точці 100 т. п. н., т. е. на фізико-генетичної карті цієї плазміди має координати 66,7-100 F (pnc. 5.36).
Залежно від контрольованих ними функцій / гд-гени плазміди F класифікують на кілька груп. Гени однієї з цих груп кодують білки, необхідні для синтезу на поверхні клітин Е. coli, містять плазміду, специфічних ниткоподібних структур (ворсинок), які отримали назву F-пілей (від англ. РШ - фимбрии), або «статевих» пілей, в кількості 1-3 на одну клітку. Такі структури можуть бути легко диференційовані на підставі їх здатності адсорбувати «статеві» бактеріофаги, які можуть лизировать лише клітини, які є господарями відповідних кон'югатівних плазмід (рис. 5.37). Роль «статевих» пілей полягає в тому, що вони забезпечують формування первинного Кон'югаційна контакту клітини-донора з реципієнтную кліткою.

Мал. 5.37. Електронна мікрофотографія «статевої» пили бактерії з адсорбованими бактеріофагами
Подальше скорочення пілей призводить до зближення кон'югується клітин і утворення поверхневого контакту між ними (Кон'югаційна «містка»), який необхідний для одностороннього переносу генетичного матеріалу від донора до реципієнта (рис. 5.38).
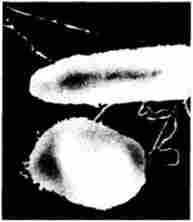
Мал. 5.38. Електронна мікрофотографія двох кон'югується бакгерій
Інші гени ГГД-області детермінують синтез білків, що забезпечують метаболізм самого процесу кон'югації бактерій і перенесення ДНК. До їх числа відносяться ферменти (ендонуклеази), які «розрізають» кільцеву молекулу плазмідної ДНК, перетворюючи її в лінійну структуру, що є обов'язковою умовою для перенесення такої молекули з клітки-донора в реципієнтную клітку.
Крім плазміди F і ряду інших плазмід, що відносяться до категорії «чистих» факторів перенесення (статевих чинників), в клітинах бактерій різних видів були виявлені кон'югатівних плазміди, що представляють собою комбіновані (коінтегратівние) структури, в яких лише одна частина є фактором переносу, а інша складається з генів , що мають хромосомні походження. Багато з таких плазмід здатні підвищувати виживаність містять їх бактерій в несприятливих умовах. Виключно широке поширення в сучасних популяціях бактерій отримали плазміди лікарської резистентності (R- плазміди), які детермінують стійкість цих організмів до антибіотиків, сульфонамідів і інших антимікробних лікарських речовин. Наявність в складі такої плазміди генів стійкості до одного або одночасно до кількох різних антибактеріальних препаратів значно ускладнює (а іноді унеможливлює) раціональне лікування інфекційних захворювань людини і тварин.
У широкому плані до плазмід резистентності відносять також ті з них, які забезпечують бактеріям стійкість до іонізуючих випромінювань, солям важких металів, бактеріофагів, бакгеріоцінам, сироватці крові та деяким іншим факторам.
Деякі плазміди можуть брати участь в контролі патогенних властивостей бактерій, впливаючи на виникнення та перебіг інфекційного процесу. Відомо, що патогенність є комплексний (полігенні) ознака, заснований на біохімічні механізми, за допомогою яких бактерії можуть викликати захворювання. Одним з важливих атрибутів патогенності є здатність бактерій прикріплятися до поверхні еукаріотів після проникнення в організм, т. Е. Здійснювати колонізацію цих клітин.
При вивченні штамів Е. coli> виділених від тварин і людей, які страждають кишковими розладами (діареєю), були виявлені різні білки (антигени колонізації), що формують на поверхні бактеріальної клітини короткі ниткоподібні вирости, названі звичайними пілямі (фімбріями). Такі фимбрии забезпечують специфічне прикріплення (адгезію) бактерії до епітеліальних клітин слизової оболонки кишечника. Поряд з хромосомними детермінантами антигенів колонізації у цих бактерій виявлені також плазміди, що кодують синтез адгезивних фімбрій. Так, в клітинах Е. coli, викликають діарею новонароджених поросят, виявлені плазміди, що детермінують синтез антигену АГ88, пов'язаного з фімбріями колонізації епітелію тонкого кишечника цих тварин. Плазміда, що кодує антиген колонізації К99, була ідентифікована у бактерій цього виду, виділених від теляти. Плазміди, що забезпечують синтез антигенів CFAI і CFAU (Від англ, colonization factor antigen - антігенфактора колонізації), виявлені в клітинах енте- ропатогенних До coli, виділених з кишечника людей з гострою діареєю. Ці антигени пов'язані з адгезивними фімбріями, необхідними для колонізації епітелію слизових оболонок людини.
Друлш найважливішим фактором патогенності бактерій є їх здатність продукувати токсини. Клітини патогенних штамів Е. coli та інших кишкових бактерій (ентеробактерій) можуть синтезувати ентеротоксини як під контролем хромосомних генів, так і під генетичним контролем плазмід, званих Ялг-плазмідами. Такі плазміди були виявлені у токсигенних бактерій, виділених від тварин і людей, які страждають на діарею. Є експериментальні дані, які показують, що включення плазмідної комплексу, що складається з двох плазмід (Ent + К88), в клітини непатогенного штаму Е. coli, виділеного від здорових поросят, перетворює ці бактерії в ентеропатогенні, що проявляється в їх здатності викликати важкі форми діареї у заражених тварин. У той же час зараження поросят аналогічними бактеріями, що містять лише одну з цих плазмід (Я88 або Ent), не приводило до розвитку захворювання або супроводжувалося проявом легких форм діареї.
Особливу категорію позаклітинних токсичних білків представляють собою гемолізіни, синтезовані різними бактеріями і здатні руйнувати мембрани еритроцитів тварин і людини. Гемолітична активність бактерій, найбільш вивчена у клітин?. coli, може контролюватися як хромосомними генами hly, так і плазмідами (Я7у-плазмідами). В даний час такі плазміди відомі у багатьох штамів цих бактерій, виділених з кишечника тварин і людини.
Численні експериментальні дані свідчать про можливість ефективного перенесення різних кон'югатівних коінте- гратівних плазмід (R, Ent, Hly, CFA і ін.) з клітин-донорів в реціпіент- ні клітини як в рамках одного виду, так і між бактеріями різної видової приналежності при їх спільному культивуванні. Отже, можливий як внутрішньовидової, так і міжвидової (межродовой) кон'югаціон- ний перенесення бактеріальних плазмід. Під час Кон'югаційна перенесення відбувається реплікація плазмідної ДНК, в результаті якої одна копія плазміди передається реципієнту, а інша залишається в клітці-до норі.
Слід зазначити, що крім великого числа кон'югатівних плазмід в клітинах різних бактерій виявлені також некон'югатівние (нетрансміссівние) плазміди (R, Ent, Hly та ін.). Вважають, що некон'югатівние плазміди природного походження позбавлені фактора перенесення (генетичної області tra) або цей фактор дефектний по своєї основної функції, тому вони не здатні забезпечувати бактеріям властивості донорів генетичного матеріалу і не можуть самостійно передаватися від одних клітин до інших. Однак їх кон'югаційний перенесення забезпечується «чистими» факторами перенесення або кон'югатівних коінтегратіінимі плазмидами, що знаходяться разом з ними в одній і тій же бактеріальної клітці. плазмида F і деякі інші кон'югатівних плазміди крім власного перенесення здатні також мобілізувати на перенесення хромосомні гени клітин, в яких вони знаходяться, якщо відбувається інтеграція такої плазміди в структуру бактеріальної хромосоми.
Таким чином, необхідно відзначити, що є значний прогрес у вивченні дії плазмід і їх впливу на патогенність бактерій. Лікарська резистентність бактерій в основному визначається плазмидами, які відносно швидко копіюються і поширюються в різних бактеріальних спільнотах в умовах селективного дії відповідних антибактеріальних препаратів. Хоча в кодуванні резистентності беруть участь і хромосомні гени бактерій, їх внесок набагато менш значний, оскільки ефективність кон'югаціонной передачі таких генів від одних бактерій до інших істотно нижче, ніж в разі генів /? - плазмід.