| Головна |
| «« | ЗМІСТ | »» |
|---|
Генетичний матеріал еукаріот має складну надмолекулярну організацію, яка визначає формування хромосом. Крім того, в клітинах різних організмів виявлені екстрахромосомние (внеядерние, цитоплазматичні) молекули ДНК.
Генетичний матеріал зрілої вірусної частки (віріона) представлений однією молекулою нуклеїнової кислоти (ДНК або РНК), яка оточена захисною білковою оболонкою (капсидом). Поряд з вірусами, що містять двухцепочечную або одноцепочечную молекулу ДНК в замкнутій (кільцевій) формі, є представники, у яких ця молекула є незамкненою (лінійної) структурою і також може бути двухцепочечной або одноцепочечной. У разі РНК-содсржащіх вірусів відомі як одноцепочечниє, так і дволанцюжкові варіанти молекул РНК. Розміри хромосом таких вірусів значно менше, ніж у більшості ДНК-co тримають вірусів, і найчастіше за все варіюють в межах від 3 000 до 7 000 рибонуклеотидов.
Молекули вірусних ДНК як правило мають довжину від 0,5 до 100 мікрометрів (мкм), тоді як довжина віріонів зазвичай коливається від 0,02 до 0,3 мкм. Настільки явна невідповідність між розмірами генетичного матеріалу і вміщує його білкового капсида усувається шляхом багаторазового закручування молекули ДНК навколо осі її спіралі, що призводить до утворення значно коротшою за довжиною суперспіралізованной (суперзакрученной) структури, яка формує хромосому вірусу (рис. 5.32).
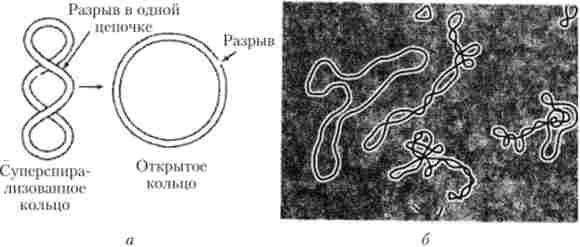
Мал. 5.32. Супсрспіралізованная молекула ДНК: а - принцип будови суперзакрученной молекули ДНК, перетворюється в відкриту кільцеву форму при розриві одного ланцюжка молекули; б - електронна мікрофотографія відкритою і закритих молекул ДНК бактеріофага
В результаті утворення супервитки лінійні розміри вірусної хромосоми виявляються в кілька сотень разів менше, ніж розміри складової її ДНК. Це наочно демонструє електронно-мікроскопічне зображення бактеріофага (фага) Т2 після його послідовної обробки гіпертонічним сольовим розчином і дистильованою водою (осмотичного шоку) (рис. 5.33). Така обробка привела до розриву білкового капсида фага і звільнення його молекули ДНК, яка раніше була щільно упакована в хромосомну структуру. Виміряна довжина цієї молекули склала близько 50 мкм, що більш ніж в 500 разів перевищує розміри фаговой головки, в якій вона перебувала.

Мал. 5.33. Електронна мікрофотографія молекули ДНК бактеріофага Т2, підданого дії осмотичного шоку
Вельми незначні розміри геномів дрібних вірусів дозволяють їм кодувати лише поодинокі білки, синтезовані метаболічної системою клітини-господаря, в яку проник генетичний матеріал вірусу.
Так, наприклад, в РНК найдрібнішого з відомих вірусів, який виділяється при некрозі листя тютюну, яка складається из 1 200 рибонуклеотидов, був виявлений лише один структурний ген, що кодує білок оболонки цього вірусу. Разом з тим ДНК дрібного фага рХ174 (5,4 х 103 пар нуклеотидів) завдяки наявності в ній перекриваються генів кодує 9 різних білків, які забезпечують формування зрілих фагових частинок. З іншого боку, хромосомна ДНК досить великого фага Т4 (1,8 х 105 пар нуклеотидів) могла б містити не менше 100 структурних генів середнього розміру (ідентифіковано більше 40 білків цього фага, синтезуються зараженими клітинами Е. з І).
Віруси володіють специфічністю відносно клітин організмів- господарів (бактерій, рослин, тварин, людини), в яких вони можуть розмножуватися. Після проникнення генетичного матеріалу вірусу в клітину в ній починається процес синтезу вірусних білків і нуклеїнової кислоти на основі генетичної програми цього вірусу і за допомогою метаболічної системи господаря. При цьому проникла РНК ретровірусів служить матрицею для синтезу комплементарної вірусної ДНК за механізмом зворотної транскрипції, тоді як у інших РНК-вірусів може відбуватися лише копіювання їх первинного генетичного матеріалу (молекул РНК).
Можливі два основні варіанти розвитку вірусу в клітці-хазяїні, які в разі бактеріальних фагів прийнято називати литичним н лізогенним циклами (Рис. 5.34). Розвиток вірулентних вірусів (фагів) зазвичай відбувається тільки по литичному шляху, пов'язаному з синтезом компонентів вірусних частинок (білків, нуклеїнових кислот), складанням ви- Ріоні і подальшим руйнуванням (лізисом) клітини-господаря. Однак для помірних (латентних) вірусів, прикладом яких є бактеріофаг X, можливий і другий варіант розвитку, що складається в лизогенизации клітини-господаря (рис. 5.34). При цьому ДНК вірусу інтегрується в хромосому хазяїна за механізмом сайт-специфічної рекомбінації і існує в ній у формі провируса (профага), репліціруясь і передаючись в процесі клітинного ділення дочірнім клітинам як складова частина хромосоми материнської клітини. Разом з тим під впливом ряду індукують факторів (ультрафіолет, іонізуюча радіація, різні хімічні агенти і ін.) Відбувається вихід вірусу з інтегрованого стану, т. Е. З форми провируса (профага), в автономне стан, що призводить до його розмноження і лизису клітини-господаря (рис. 5.34). Іншими словами, можливий перехід зі шляху лизогенизации на литичний шлях розвитку вірусу і назад. Аналогічні шляхи розвитку відомі і в разі ряду вірусів еукаріотів, в тому числі для ретровируса імунодефіциту людини, що є причиною виникнення СНІДу.
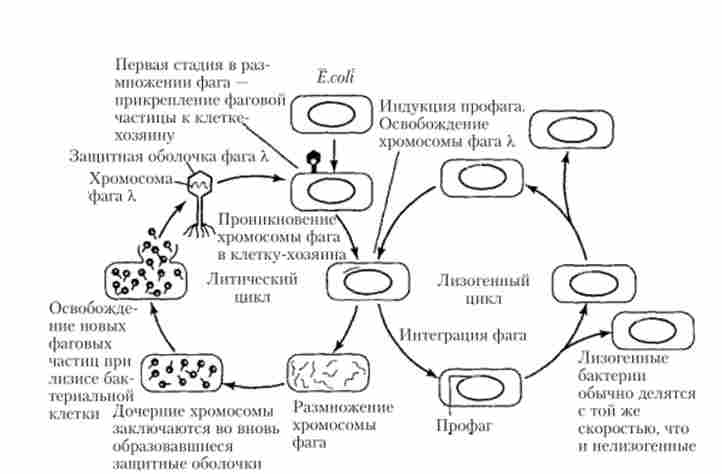
Мал. 5.34. Два варіанти можливого розвитку помірного фага X в клітинах Е. coli
На відміну від вірусів хромосома бактеріальної клітини містить молекулу ДНК лише одного типу, а саме кільцеву двухцепочечную ДНК набагато більш значних розмірів (1 000-2 000 мкм завдовжки). Оскільки у бактерій немає справжнього ядра, то їх генетичний матеріал організований у вигляді ядерноподобной структури (нуклеоида), що розташовується в цитоплазмі клітини. Кожному нуклеоида відповідає одна хромосома, т. Е. Бактерії є гаплоїдними організмами.
Як і у вірусів, лінійні розміри генетичного матеріалу бактерій явно не відповідають розмірам структурного утворення, в якому він знаходиться. Так, наприклад, типові клітини Е. coli мають форму паличок довжиною 1-5 мкм і товщиною 0,4-0,8 мкм. Ця невідповідність також усувається шляхом супере бенкет ализации молекули ДНК, яка формує хромосому нуклеоида.
В процесі упаковки молекули беруть участь кілька ДНК-зв'язуючих білків, виявлених в клітинах Е. coli, частина яких за своїм амінокислотним складом нагадують гистони з хромосоми еукаріотів.
Крім того, в нуклеоида цих бактерій знайдена РНК, роль якої залишається неясною. Пості обробки нуклеоідного комплексу Е. coli ферментами, які руйнують білки і РНК, виникають відкриті кільцеві форми хромосомної ДНК, які вивчають за допомогою електронномікроскопічних і радиобиологічних методів (рис. 5.35). При цьому можна визначити довжину молекул.

Мал. 5.35. Радіоавтограф кільцевої молекули ДНК хромосоми Е. coli. Видно ділянку почалася реплікації молекули
Розміри хромосомного генома бактерій дозволяють їм мати більш значне, ніж у вірусів, кількість структурних генів для синтезу білків, що забезпечують всі процеси життєдіяльності цих організмів. Так, наприклад, хромосомна ДНК клітин Е. coli містить 4,2 х 10б пар нуклеотидів, що було б достатнім для формування приблизно 4 000 генів середнього розміру. До теперішнього часу в єдиній кільцевої хромосомі (одній групі зчеплення) цих бактерій ідентифіковано понад 650 різних генних локусів.
У хромосомах бактерій, що викликають інфекційні захворювання тварин і людини, виявлені різні гени, що детермінують патогенні властивості цих бактерій (формування антигенних структур, синтез токсинів, здатність руйнувати еритроцити крові і ін.). Значний клінічний інтерес представляють також гени стійкості бактерій до антибіотиків та інших антибактеріальних препаратів.