| Головна |
| «« | ЗМІСТ | »» |
|---|
транслокация є реципрокний обмін ділянками хромосом. В результаті такого обміну у гомозигот по транслокаціях змінюється характер зчеплення генів. У гетерозиготі по транслокації гени, що належать до різних, що не гомологічним хромосомами, успадковуються як належать до однієї групи зчеплення. Це пояснюється тим, що повністю функціональними виявляються тільки ті суперечки (у рослин) і гамети (у тварин), які несуть батьківські поєднання хромосом.
Характер кон'югації транслоцироваться хромосом змінюється: утворюється фігура хреста. Щільна кон'югація поблизу точок розривів виявляється утрудненою, що призводить до пригнічення кросинговеру в цих ділянках.
У гетерозиготи по транслокації в профазі мейозу утворюються квад- ріваленти, а не біваленти, як зазвичай, оскільки гомологічні ділянки виявляються у всіх чотирьох кон'югується хромосом. З шести можливих типів гаплоїдних продуктів, що виникають при трьох способах розбіжності хромосом, тільки два типи функціонують нормально - ті, які отримують повні набори генів, характерні для вихідних батьківських форм (рис. 9.6). Решта чотири типи несуть дуплікації і браку, тому як правило не дають життєздатного потомства або не беруть участь в заплідненні.
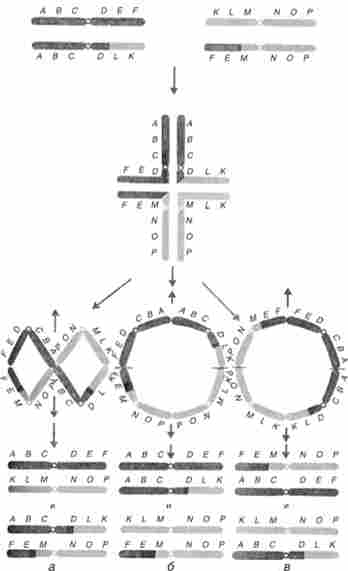
Мал. 9.6. Мейоз у гетерозиготи по реципрокною транслокации: а - повноцінні тільки два типи продуктів мейозу;
6, в- чотири типи гаплоїдних продуктів несуть делеции і дуплікації
Г'етерозіготи по реципрокним транслокаціях у тварин зустрічаються рідко, але широко поширені у рослин. Характерний приклад в цьому відношенні представляють різні види ослінніка (Oenothera). Наприклад, у О. lamarkiana з 14 хромосом 12 залучені в реципрокні транслокації, тому в мейозі у цієї рослини спостерігають один бивалент і мультівалент, що включає інші 12 хромосом. У інших видів ослінніка число хромосом, що утворюють мультіваленти, варіює, що відображає число реципрокних транслокацій. У О. muricata і О. biennis всі 14 хромосом утворюють один мультівалент, а пізніше, в діакінезе, в результаті терміналізації хиазм - кільце з 14 хромосом. Функціонуючими виявляються тільки ті два типи мікро- і мегаспор, які отримали повні набори плечей хромосом. Таким чином, нормальне запліднення відбувається тільки при об'єднанні типів гамет, що вносять в зиготу цілі батьківські комплекси транслоцироваться хромосом. Злиття гамет, що несуть однакові хромосомні комплекси, летально, і сумісні при заплідненні комплекси хромосом у різних видів Oenothera неоднакові. Завдяки такому «комплексному» запліднення види ослінніка підтримують збалансовану гетерозиготность по транслокаціях навіть при самозапилення. Рідкісні порушення сумісних комплексів хромосом були прийняті Г. де Фріз за мутації.
Подібно інверсія, транслокації забезпечують ізоляцію нових форм і сприяють дивергенції в межах виду. Особливий тип транслокаций, так звані Робертсонівські транслокации, або злиття, призводить до зміни числа хромосом. Якщо дві телоцентрічні хромосоми зливаються в області центромери, то утворюється одна метацентрична хромосома. Цей тип хромосомних перебудов отримав назву по імені дослідника У. Р. Робертсона, з'ясував механізм такого злиття.
Знання механізму наслідування і летального ефекту багатьох аберацій послужило основою генетичного методу боротьби з шкідливими комахами. Принцип цього методу сформулював А. С. Серебровський (1940). У природне середовище випускають велику кількість самців подавляемого виду, попередньо підданих дії іонізуючого випромінювання здійснюватиме. У сперматозоїдах таких самців виникають численні хромосомні перебудови. Опромінені самці конкурують за спаровування з природними самцями, а утворюються зиготи або зиготи, що виникли у їх нащадків, з великою частотою виявляються нежиттєздатними.
Останнім часом синтезовано деякі сполуки, переважно викликають хромосомні аберації, а не розумні мутації. До їх числа, наприклад, відноситься діепоксіоктан, який викликає у нейроспори і дріжджів переважно хромосомні перебудови. Такі мутагени застосовують в якості еталонних при вивченні генетичних ефектів забруднення навколишнього середовища.