| Головна |
| «« | ЗМІСТ | »» |
|---|
Згідно з уявленнями, що склалися в рамках класичної (формальної) генетики, геноми різних організмів характеризуються значною стабільністю і піддані лише дуже повільним еволюційним перетворенням. Ці уявлення істотно змінилися в зв'язку з відкриттям транспозіруемих (мігруючих) генетичних елементів (фрагментів ДНК), які можуть переміщатися як у межах одного генома, так і з одного генома в інший. Здатність до транспозиції закодована в нуклеотидних послідовностях самих мігруючих фрагментів ДНК і пов'язана з можливістю їх включення (інтеграції) в різні місця (сайти) хромосом і позахромосомних молекул ДНК і виходу ( «вирізання») з цих сайтів. Такі мобільні структури були виявлені в геномах як прокариот, так і еукаріот.
У бактерій транспозіруемие генетичні елементи представлені сегментами ДНК двох типів: инсерционно (вставними) послідовностями, або / 5-елементами (від англ, insertion sequences - вставні послідовності), і транспозонами. Одні й другі можуть переміщатися в межах однієї генетичної структури (бактеріальної хромосоми, плазміди) або з однієї структури в іншу (наприклад, з хромосоми в плазміду, з плазміди в хромосому, з однієї плазміди в іншу, з плазміди бактерії одного виду в плазміду або хромосому бактерії іншого виду і т. д.).
/ 5-елементи - це невеликі ділянки ДНК, розміри яких зазвичай варіюють в межах від 760 до 2 000 пар нуклеотидів. На кінцях (флангах) цих ділянок є однакові або дуже подібні короткі нуклеотидні послідовності (20-40 пар нуклеотидів), розташовані в зворотному порядку (інвертовані повтори). Вважають, що кінцеві повтори (фланкирующие послідовності) відіграють важливу ю роль в процесах переміщення транспозіруемих елементів, хоча точний механізм таких переміщень недостатньо вивчений.
Різні / 5-елемснти є типовими «мешканцями» бактерій і зазвичай виявляються в кількостях, що перевищують одну 'копію на клітку (табл. 5.6). Наявність ідентичних / 5-елементів в різних бактеріальних структурах (хромосомах, плазмидах) створює основу для їх взаємодії за механізмом сайт-специфічної рекомбінації, що може призводити до об'єднання (коінтеграції) таких структур або до генетичного обміну між ними. На відміну від генетичної рекомбінації загального типу, що вимагає наявності значних областей гомології ДНК у двох взаємодіючих родинних структур, сайт-специфічна рекомбінація заснована на тому, що відповідний фермент дізнається невеликі специфічні послідовності в двох місцях (сайтах) однієї або різних молекул ДНК, де і відбувається процес рекомбінації. Отже, така рекомбінація може призводити до генетичного обміну між несумісними (негомоло- гічної) структурами (хромосомами, плазмидами).
Таблиця 5.6
Зміст инсерционно послідовностей в хромосомному геномі Е. сон
|
Назва |
Число копій в клітці |
Розміри однієї копії |
|
послідовності |
на хромосому |
(В парах нуклеотидів) |
|
/ 51 |
5-8 |
768 |
|
/ 52 |
5 |
1 327 |
|
/ 53 |
5 |
1 400 |
|
/ 54 |
1-2 |
1 400 |
Транспозони є більш складними утвореннями з розмірами порядку 2 000-20 000 пар нуклеотидів, в серединній частині яких знаходяться ті чи інші функціонально значущі гени, а кінцеві ділянки містять / S'-елементи або фланковані іншими довгими інвертованими або однаковими (прямими) нуклеотидними повторами ( 800-1 500 пар нуклеотидів). У складі деяких транспозони виявлені гени, що кодують синтез ферментів, названих транспозази і резолвазалш, які вважаються відповідальними за процеси транспозірованія. Разом з тим транспозони можуть містити гени, що не мають відношення до транспозиції, але які відіграють важливу роль у виживанні бактерій при несприятливих умовах середовища проживання (наприклад, гени стійкості до антибіотиків та інших антибактеріальних препаратів). Такі гени переміщаються разом з усім транспозони з однієї бактеріальної плазміди в іншу, з плазміди в хромосому ит. д.
Вважають, що специфічні кінцеві повтори / S-елементів та транспозони грають ключову роль в процесах транспозиції, оскільки вони розпізнаються ферментами, які здійснюють сайт-специфічну рекомбінацію. Разом з тим в разі ряду транспозони отримані експериментальні дані, які свідчать про те, що при транспозиції може працювати механізм, який не пов'язаний з процесом повного «вирізання» всієї структури з одного сайту і її переміщення в інший сайт (сайт-мішень). При цьому відбувається подвоєння (реплікація) генетичного елемента, одна копія якого залишається в вихідному сайті, а інша переміщається в новий сайт-мішень. Іншими словами, така транспозиція полягає в збільшенні числа копій відповідного транспозона. Якщо ж все-таки відбувається виключення ( «вирізування») транспозона, то цей процес не завжди буває достатньо точним і може призводити до збереження в структурі ДНК колишньої мішені тих чи інших залишків послідовностей транспозіруемого елемента. Відомо також, що вбудовується транспозон або / S-елемснт фланкірується короткими повторами ДНК господаря (4-12 пар нуклеотидів). Є підстави вважати, що транспозіруе- мие елементи грають важливу роль в процесах еволюції мікроорганізмів, забезпечуючи специфічну форму їх внутрішньовидової і міжвидової генетичного обміну (так званий горизонтальний перенос генів). Відомо також, що при інтеграції мобільних структур до відповідних сайти хромосом і плазмід і при їх «вирізання» в цих та суміжних сайтах можуть відбуватися мутаційні перебудови генетичного матеріалу (інверсії, делеції, транслокації та ін.), Т. Е. Транспозіруемие елементи в цьому випадку виступають в ролі біологічних мутагенів.
Мігруючі генетичні елементи еукаріот, вперше виявлені в 1951 р у вищих рослин (кукурудзи), були потім виявлені і в інших організмів. Зміст деяких з цих елементів в еукаріотичних геномі може бути досить значним, т. Е. Вони є багато- копийности нуклеотидними послідовностями ДНК. Разом з тим функціональне значення таких послідовностей залишається багато в чому незрозумілим.
Мобільні структури невеликих розмірів, що містять кінцеві повтори і найбільш подібні до транспозонами бактерій, були виявлені в різних ділянках геномів дріжджів і дрозофіли. До числа найбільш вивчених мігруючих елементів дрозофіли відноситься група близьких за будовою нуклеотиднихпослідовностей, позначених загальним терміном copia, які в сукупності складають значну частину геномної ДНК цих організмів.
У різних ділянках генома людини виявляється багаторазово повторюється (близько 500 тис. Копій на гаплоїдний геном) нуклеотидних послідовність, що складається з 300 пар нуклеотидів, яка отримала назву Л / и-послідовності. Оскільки кінцеві ділянки цієї послідовності фланковані короткими прямими повторами (7-20 пар нуклеотидів), що визначають її схожість з відомими транспозонами, то вважають, що вона також відноситься до категорії мобільних елементів еукаріот. Подібні Л / и-подібні послідовності виявлені і в геномах різних ссавців.
Як джерело мобільних структур прокариотичного і еукаріотичного генома можна розглядати також геномної ДНК різних вірусів, які здатні інтегруватися в ті чи інші ділянки ДНК бактерій і еукаріот і деякий час існувати там в формі провируса, т. Е. Як складова частина генетичної структури клітини господаря . Прикладом є інтеграція бактеріофага X в хромосомну структуру клітини Е. coli за допомогою механізму сайт-специфічної рекомбінації (рис. 5.31). Ця особливість стосується і РНК-містять ретровірусів, ДНК яких, синтезована на основі розглянутого вище механізму зворотної транскрипції, включається в хромосомну структуру еукаріотичної клітини і потім може довго реплицироваться в її складі, передаючись від материнської клітини до дочірнім в процесі мітотичного поділу. При подальшому неправильному (неточному) «вирізання» з генома господаря вірус залишає в ньому ті чи інші фрагменти власного геному і набуває окремі ділянки генетичного матеріалу хазяїна.
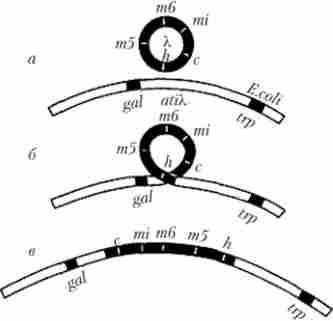
Мал. 5.31. Інтеграція бактеріофага X в хромосомний сегмент клітини Е. coli між локусами генів gal (Утилізації галактози) і trp (Синтез триптофану): а - кільцева хромосома фага X (m5, m6, mi, c, h- окремі гени фаговой ДІК) і фрагмент кільцевої хромосоми Е. coli (att Х-сайт гомології з фагової ДНК); б - сайт-специфічна рекомбінація, пов'язана з розривами хромосоми фага між локусами Лісі бактеріальної хромосоми між локу самі gal і trp; в - бактеріальна хромосома, яка містить ДНК фага X в формі профага
Можна вважати, що результатом зазначених подій є генетичні елементи клітин тварин і людини, названі резроелеменгамі (рстротранспозонамі), здатні до переміщень завдяки наявності у них коротких кінцевих повторів. Такі ретротранспозонов можуть, зокрема, кодувати білки, необхідні для здійснення в еукаріотичної клітці процесу зворотної транскрипції PHК-містять ретровірусів, які розглядаються в якості причини виникнення злоякісних пухлин (онкологічних захворювань) і синдрому набутого імунодефіциту (СНІДу). Так, отримані експериментальні дані, які свідчать про те, що деякі онкогени клітин людини, пов'язані з появою злоякісних новоутворень, мають структурну гомологію з РНК ретровірусів.
Значення транспозіруемих елементів еукаріот пов'язують насамперед з їх мутагенну дію, а також з імовірним участю в процесах генетичної регуляції, оскільки, інтегруючись по сусідству з деякими генами, вони можуть робити істотний вплив на їх експресію. Стає очевидною також роль ретротранспозонов, представлених нуклеотидними послідовностями деяких ретровірусів, у розвитку ряду захворювань тварин і людини (лейкози птахів і ссавців, злоякісні новоутворення, СНІД і ін.). Слід ще раз підкреслити ймовірну роль мігруючих елементів як одного з факторів еволюції організмів.
Таким чином, експериментальні досягнення, які привели до розшифровки структури генетичного коду і встановлення механізмів його реалізації, не тільки відкрили нові можливості розуміння живої природи, а й забезпечили рішення ряду практичних завдань на основі маніпуляцій з молекулами ДНК з організмів різних видів (успіхи в галузі генетичної інженерії, біотехнології та ін.). Ці досягнення стали також основою для розробки нових напрямків у медичній і клінічної генетики, пов'язаних з діагностикою та лікуванням спадкової патології людини (генна діагностика, генна терапія і ін.).