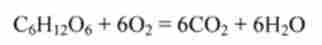
| Головна |
| «« | ЗМІСТ | »» |
|---|
Биоорганичні речовини, такі, як глюкоза, володіють великим запасом енергії. При окисленні глюкози киснем
вивільняється енергія Гіббса AG = -2880 кДж / моль. Ця енергія може запасатися в клітці у формі хімічної енергії фосфатних зв'язків аденозілтріфос- фата АТР. Утворені молекули АТР дифундують в різні ділянки клітини, де використовується енергія. АТР - це переносник енергії. Клітина використовує цю енергію для виконання роботи. Однак при гліколізі витрачається лише незначна частина енергії, запасеної в глюкозі (кілька відсотків). Основна її частина передається в циклі Кребса (рис. 9.4), зв'язаному з клітинним диханням.
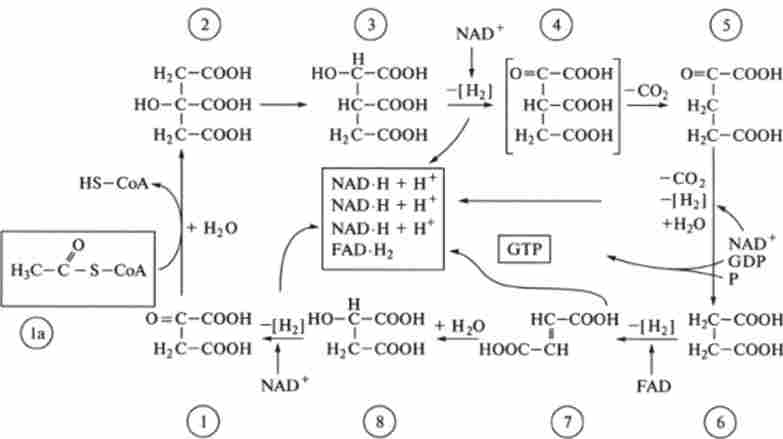
Мал. 9.4. Цикл Кребса (цикл лимонної кислоти)
I - оксалоацетат, 1а ацетил * Сол, 2 - лимонна кислота (цитрат). 3 - іеоцітрат. 4 - оксалосукцінат. 5 - кетоглугарат. 6 - бурштинова кислота (сукцинат). 7 - фумарат. 8 - яблучна кислота (малат)
Цикл Кребса, або цикл лимонної кислоти, або цикл 3-карбонових кислот, являє собою ряд послідовних реакцій, що протікають в мітохондріях. В ході цих реакцій здійснюється катаболізм ацетильних груп СН3СО-, переданих від пірувату, кінцевого продукту гліколізу. Піруват вступає в реакції циклу Кребса, попередньо перетворюючись в ацетил-СоА.
Цикл Кребса, як і гліколіз, являє собою метаболічний шлях, що складається з послідовних стадій - реакцій. На відміну від гліколізу, цей шлях замкнутий, циклічний.
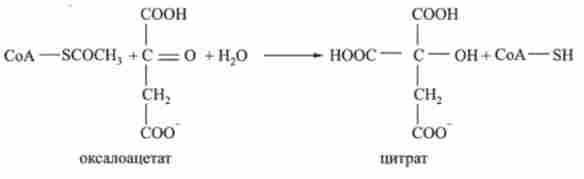
1. Ацетил-СоА - продукт катаболізму вуглеводів, білків і ліпідів - вступає в цикл, реагуючи (конденсується) з сіллю щавелевоуксусной кислоти (оксало- ацстатом). При цьому утворюється сіль лимонної кислоти (цитрат):
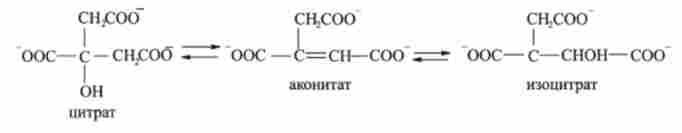
2. Цитрат изомеризуется в ізоцитрат. Реакція каталізується ферментом ако- нітазой і проходить через освіту аконітата з подальшим його перетворенням в ізоцитрат:
3. ізоцитрат окислюється до а-кетоглутарата. Реакція каталізується ферментом ізоцитратдегідрогеназа:
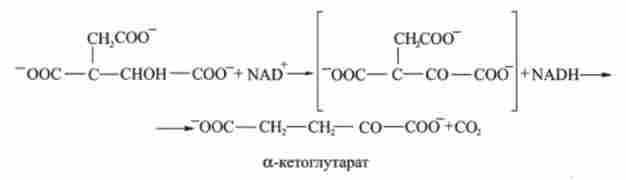
4. а-кетоглутаратом піддається окислювальному декарбоксилюванню з утворенням сукцинил-СоА. Каталізує а-кетоглутарат дегідрогеназ:
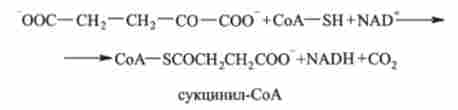
5. сукцініл-СоА перетворюється в сукцинат. Реакція каталізується ферментом сукцинат-СоА-лігази:
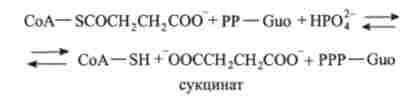
6. Сукцинат перетворюється в фумарат. Реакція каталізується ферментом дегідрогеназ:
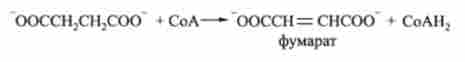
7. Фумарат гідратіруется по подвійному зв'язку з утворенням малата (сіль яблучної кислоти). Каталізується фумаратгідратазой:
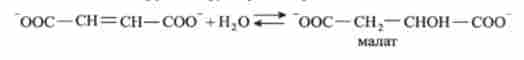
8. Манат окислюється до оксапоацетата. Каталізується мапатдегідрогеназой:

Мал. 9.5. Будова мітохондрій (збільшення приблизно 25 000)
На восьмий стадії цикл замикається і починається його нове проходження.
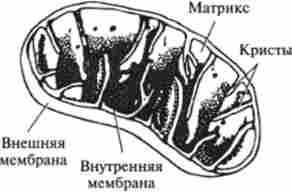
Всі стадії циклу лимонної кислоти протікають у внутрішньому середовищі мітохондрій - матриксе (рис. 9.5). Тут знаходяться всі ферменти цього метаболічного шляху.
Мітохондрія (від грец. «Mitos» - нитка і «chondrion» - зернятко) має витягнуту форму; довжина 1,5-2 мкм, діаметр 0,5-1 мкм. Органели клітин тварин знаходяться в рідкому середовищі клітини - цитоплазмі (див. Рис. 6.2).
Внутрішній простір мітохондрій оточене двома безперервними мембранами. При цьому зовнішня мембрана гладка, а внутрішня утворює численні складки, або Крісті. Внутрімітохондріальное простір обмежений внутрішньою мембраною, заповнене рідким середовищем - матриксом, який приблизно на 50% складається з білка і має дуже тонку структуру. Подовжена форма мітохондрій не універсальна. У деяких тканинах, наприклад в поперечно-смугастих скелетних м'язах, мітохондрії іноді набувають химерних обрисів.
В мітохондріях зосереджена велика кількість ферментів.
У клітці може перебувати від кількох сотень до кількох десятків тисяч мітохондрій. Для одного і того ж типу клітин число мітохондрій більш-менш постійно. Однак слід пам'ятати, що кількість мітохондрій може змінюватися в залежності від стадії розвитку клітини і її функціональної активності, а в цілому від інтенсивності навантажень на організм.
Мітохондрії - енергетичні станції, що виробляють енергію для життєдіяльності організму. Особливо багато мітохондрій в м'язових клітинах, де потрібні великі витрати енергії.
Освічені в циклі Кребса високоенергетичні речовини NADH і FADFb (див. Рис. 9.4) передають свою енергію в реакції ресинтезу АТР з ADP:
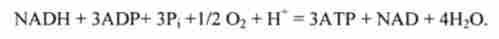
В результаті на кожну молекулу NADH утворюються 3 молекули АТР. Ця реакція окислювально-відновна, т. Е. Супроводжується перенесенням електронів від відновника NADH до окислювача (див. Розд. 4.3). В якості окислювача виступає кисень О2. Ця реакція називається окислювальним фосфорилюванням ADP в АТР.
Окислювальне фосфорилювання відбувається у внутрішній мітохондріальній мембрані. У трьох ділянках дихального ланцюга запасається енергія в результаті синтезу АТР з ADP і Р ,.
Реакція протікає в кілька стадій на внутрішніх мембранах мітохондрій (див. Рис. 9.5), в системі ферментів, званої дихальної ланцюгом. Сюди з клітинної плазми надходять молекули ADP. Відповідний окіслітельновосстановітельние процес називається клітинним диханням. Саме тут витрачається кисень, яким ми дихаємо.
Молекули АТР, утворені в матриксі, виходять з мітохондрій в плазму клітини, де беруть участь у різних біохімічних реакціях, що протікають з витратою енергії.
Таким чином, енергія, що вивільняється в процесі перенесення електронів від відновників, використовується для окисного фосфорилювання ADP в АТР.
Припускають, що енергія, що вивільняється вдихательная ланцюга, витрачається безпосередньо на переклад внутрішньої мембрани в нове, багате енергією конформационное стан, яке, в свою чергу, стає рушійною силою окисного фосфорилювання, що призводить до утворення АТР. В даний час найбільш серйозне обґрунтування отримала гіпотеза хемоосмо- тичного сполучення Мітчела.
Таким чином, біосинтез АТР в тваринному організмі здійснюється з ADP і неорганічного фосфату Р, при активації останнього за рахунок енергії окислення органічних сполук при метаболічних процесах.
Окислення органічних сполук в живих системах не завжди пов'язане з фосфорилюванням, і фосфорилювання не обов'язково має бути окислювальним.
Відомо кілька сотень реакцій окислення. Не менш десятка з них пов'язане з одночасним активированием неорганічного фосфату. Такі реакції називають реакціями субстратного фосфорилювання. Тут реакції розщеплення субстрату супроводжуються передачею енергії безпосередньо неорганичному фосфату. В результаті утворюється інший фосфорілірованний субстрат з макроергічним зв'язком. В цьому випадку в процесі не бере дихальна ланцюг ферментів і не відбувається перетворення енергії, що виділяється при перенесенні електронів на кисень, в енергію фосфатного зв'язку АТР.
Як приклад субстратного фосфорилювання можна привести реакцію перетворення сукніціл-СоА в бурштинову кислоту з утворенням GTP з GDP і фосфату Р, в лимоннокисле циклі.
У рослинах джерелом енергії для активування неорганічного фосфату і забезпечення синтезу АТР служить енергія сонячного світла, вловлюється фото- синтетичним апаратом клітини. Таке фосфорилювання називають фотосін- тетичного.
Для задоволення потреб людського організму в енергії молекули АТР протягом доби тисячі і тисячі разів розщеплюються до молекул ADP і Р, з подальшим ресинтезом АТР. Крім того, швидкість ресинтезу АТР повинна змінюватися в широких межах - від мінімальної під час сну до максимальної в періоди напруженої м'язової роботи.
Зі сказаного можна зробити висновок, що окисне фосфорилювання не просто безперервний життєво важливий процес. Він повинен регулюватися в широких межах, що досягається шляхом тренування.
Сумарне рівняння реакцій гліколізу і циклу лимонної кислоти записується в такий спосіб:
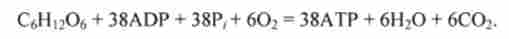
Стандартна енергія Гіббса окислення 1 моля глюкози СБН ^ Про дорівнює ДG * = = -2880 кДж (см. Розд. 5.1). Стандартна енергія Гіббса гідролізу 38 молей АТР (запасені енергія) дорівнює ДG ° ' = -38 * 30 = -1180 кДж, т. Е. Запасається лише 40% енергії глюкози (коефіцієнт корисної дії дихання). Інша енергія виділяється з організму у вигляді тепла Q. Цим пояснюється розігрів і підвищення температу ри тіла при інтенсивній роботі (див. Рис. 5.2).
Глюкоза виконує функцію клітинного палива в нашому організмі. Вона виходить головним чином або в процесі травлення з вуглеводів, або шляхом синтезу з резервних жирів.