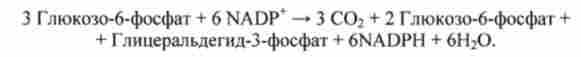
| Головна |
| «« | ЗМІСТ | »» |
|---|
О. Варбург в ході з'ясування механізму гліколізу виявив в червоних кров'яних клітинах - еритроцитах - новий фермент. Цей фермент, названий проміжним ферментом (Zwischenferment), каталізує окислення глюкози, але не входить в шлях гліколізу.
Подальші дослідження дозволили виявити новий шлях розщеплення глюкози, названий пентозофосфатний шляхом (циклом) або гексозомонофосфатного шунтом.
Сутність шляху була встановлена, коли визначили природу проміжного ферменту Варбурга. Виявилося, що цим ферментом є глюкозо-6-фосфат дегидрогеназа. Коферментом для неї служить NADP.
Пентозофосфатний цикл не призводить до синтезу АТР, він виконує дві основні функції: 1) освіту NADPH для відновлювальних синтезів, таких, як синтез жирних кислот і стероїдів; 2) забезпечення рибозой синтезу нуклеотидів і нуклеїнових кислот.
Пентозофосфатний шлях є альтернативним шляхом окислення глюкози. Цей обхідний шлях (шунт) включає 8 стадій:
На 8-й, заключній стадії пентозофосфатного шляху фосфогексоізомераза перетворює дві молекули фруктозо-6-фосфату в глюкозо-6-фосфат. На цій стадії пентозофосфатний шлях перетинається з шляхом гліколізу (шунт замикається).
В результаті з трьох молекул глюкозо-6-фосфату утворюються три молекули СОГ і три молекули пентоз - рибози і ксилози. Вони використовуються для регенерації двох молекул глюкозо-6-фосфату і однієї молекули глицеральдегид-3- фосфату. Особливо важливе значення має освіту рибози, яка необхідна для синтезу нуклеїнових кислот.
Оскільки з двох молекул глицеральдегид-3-фосфату можна регенерувати молекулу глюкозо-6-фосфату, глюкоза може бути повністю окислена при перетворенні по пентозофосфатному шляху:
Пентозофосфатний шлях істотно відрізняється від гліколізу: на першій стадії здійснюється дегидрирование (окислення) глюкозо-6-фосфату. В якості окислювача виступає не NAD, як в гліколізі, a NADP. Одним з продуктів пентозофосфатного шляху є вуглекислий газ СО2, який в реакціях гліколізу не утворюється. Нарешті, пентозофосфатний шлях не генерує АТР.
Пентозофосфатний шлях протікає в печінці, жировій тканині, корі надниркових залоз, щитовидної залозі, еритроцитах, насінниках і в молочних залозах в період лактації. Цей шлях метаболізму вуглеводів не діє в нелактірующіх молочних залозах і малоактивний в скелетних м'язах.
Всі тканини, в яких активність даного шляху висока, використовують в реакціях відновного синтезу NADPH, наприклад в реакціях синтезу жирних кислот, стероїдних гормонів, амінокислот (за участю глутаматдегідрогенази) або відновленого глутатіону в еритроцитах.
При інтенсивному синтезі жирів (Ліпогенез) і при функціонуванні систем за участю NADPH зростає швидкість деградації глюкози по пентозофосфатному шляху. Це пов'язано зі збільшенням відносини концентрацій NADP / NADPH. Після прийому їжі може індукувати синтез глюкозо-6 фосфатдегідрогенази і 6-фосфоглюконатдегідрогенази.
Рибоза з пентозофосфатного шляху надходить в систему синтезу нуклеотидів і нуклеїнових кислот. Джерелом рибози є интермедиат рибоза-5-фосфат.
М'язова тканина містить малі кількості глюкозо-6-фосфатдегідрогенази і 6-фосфоглюконатдегідрогенази. Проте скелетний м'яз здатна синтезувати рибозу. Синтез, мабуть, здійснюється при зверненні неокислювального фази пентозофосфатного шляху, що утилізує фруктозо-6-фосфат.
Таким чином, синтез рибози може здійснюватися в тканини, якщо в ній протікає частина реакцій пентозофосфатного шляху.