| Головна |
| «« | ЗМІСТ | »» |
|---|
ембріональна індукція - це взаємодія частин зародка, при якому одна ділянка зародка впливає на долю іншої ділянки. Явище ембріональної індукції з початку XX ст. вивчає експериментальна ембріологія.
Класичними вважають досліди німецького вченого Г. Шпемана і його співробітників (1924) на зародках амфібій. Для того щоб мати можливість простежити за долею клітин певної ділянки зародка, Шпеман використовував два види тритонів: тритона гребінчастого, яйця якого позбавлені пігменту і тому мають білий колір, і тритона смугастого, яйця якого завдяки пігменту мають жовто-сірий колір.
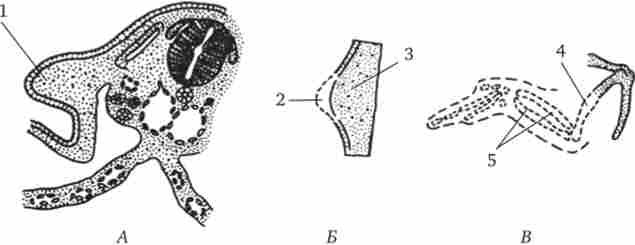
Один з дослідів полягає в наступному: шматочок зародка з області дорсальній губи бластопора на стадії гаструли тритона гребінчастого пересаджують на бічну або вентральній сторону гаструли тритона смугастого (рис. 8.8). У місці пересадки відбувається розвиток нервової трубки, хорди і інших органів. розвиток
може досягти досить просунутих стадій з утворенням додаткового зародка на бічній або вентральної стороні зародка реципієнта. Додатковий зародок містить в основному клітини зародка реципієнта, але світлі клітини зародка-донора теж виявляються в складі різних органів.
З цього і подібних дослідів слід кілька висновків. По перше, ділянку, взятий з спинний губи бластопора, здатний направляти або навіть перемикати розвиток того матеріалу, який знаходиться навколо нього, на певний шлях розвитку. Він ніби організовує, або індукує, розвиток зародка як в звичайному, так і в нетиповому місці. По-друге, бічна і черевна сторони гаструли володіють більш широкими потенціями до розвитку, ніж їх пре- зумптівное (передбачуване) проспективне напрямок, так як замість звичайної поверхні тіла в умовах експерименту там утворюється цілий зародок. По-третє, досить точну будову новостворених органів в місці пересадки вказує на ембріональну регуляцію. Це означає, що фактор цілісності організму призводить до досягнення хорошого кінцевого результату з нетипових клітин в нетиповому місці, як би керуючи процесом, регулюючи його в цілях досягнення цього результату.
Г. Шпеман назвав спинну губу бластопора первинним ембріональним організатором. Первинним тому, що на більш ранніх стадіях розвитку подібних впливів виявити не вдавалося, а організатором тому, що вплив відбувалося саме на морфогенез. В даний час встановлено, що головна роль в спинний губі бластопора належить хордомезодермальному зачатки, який назвали первинним ембріональним індуктором, а саме явище, при якому одна ділянка зародка впливає на долі іншого, - ембріональної індукцією.
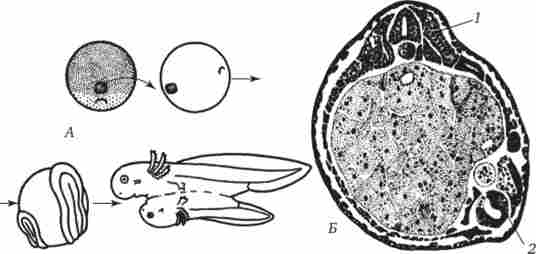
Мал. 8.8. Пересадка спинний губи бластопора від зародка-донора на черевну сторону зародка реципієнта. А - схема досвіду; Б - поперечний зріз на стадії закладки двох комплексів осьових органів.
У 30-і рр. дослідники намагалися встановити природу індукуючогоефекту. Незабаром з'ясувалося, що різноманітні убиті тканини, витяжки з найрізноманітніших тканин безхребетних і хребетних тварин, а також рослин, кілька класів хімічних сполук (білки, нуклеопротеїнами, стероїди і навіть неорганічні речовини) можуть викликати індукцію. Таким чином була встановлена хімічна природа організаторів. Одночасно стало ясно, що специфічність відповіді прямо не пов'язана з хімічними властивостями індуктора.
Увага ембріологів переключилася на індуковані тканини. Виявилося що специфічність дії індуктора-подразника може бути дуже різною, а сам ефект індукує впливу обмежується здатністю того чи іншого ділянки зародка сприймати цей вплив і відповідати на нього.
Деякі індуктори, мабуть, більше або менш специфічні у визначенні долі индуцируемой тканини. Про це свідчать наступні досліди. Якщо пересадити спинну губу ранньої гаструли, то індукується розвиток структур переднього мозку (головний індуктор), якщо ж пересадити спинну губу пізньої гаструли, то розвиваються спинний мозок і мезодермальниє тканини (тулубовий індуктор, рис. 8.9). Було показано також, що найбільш сильне нейралізующее впливає фракція нукле- опротеінов, а мезодермалізующім індуктором виявився білок. Якщо імплантувати обидва ці індуктора у вигляді суміші клітин або суміші речовин, то виходять добре розвинені зародки.

Мал. 8.9. Результати пересадки головного (А) і туловищного (Б) індукторів. Пояснення см. В тексті
Інші індуктори діють як неспецифічні пускові механізми, як би вивільняючи відповідь, вже детермінований в клітинах индуцируемой тканини. Було показано, що, наприклад, слуховий пухирець виступає не тільки в ролі індуктора слухового апарату, а й є активатором різних морфогенетичних процесів. Будучи пересаджений в область бічній лінії ембріона тритона, він тягне за собою індукцію кінцівки. Кінцівка можна індукувати також пересадкою носової плакоди або гіпофіза. Найлегше додаткові кінцівки індукуються в області бічної лінії, але вони можуть бути отримані і на черевній стороні. Ці приклади вказують на те, що специфічний відповідь залежить не стільки від індуктора, скільки від реагує області.
Здатність ембріонального матеріалу реагувати на різного роду впливу зміною своєї презумптівного долі отримала назву компетенції. Встановлено, наприклад, що компетенція до утворення нервової системи у амфібій зачіпає всю ембріональну ектодерму і виникає з моменту початку гастру- ляции. До кінця гаструляціі ця компетенція припиняється. Таким чином, зміна ходу розвитку можливо лише в тому випадку, якщо область компетенції до утворення деякої закладки ширше, ніж область, з якої вона в нормі розвивається, а також якщо індукційне дія відбувається в певний інтервал онтогенетичного розвитку.
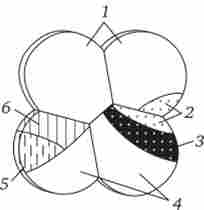
Мал. 8.10. Карта презум- птівних зачатків у зародка асцидій на стадії восьми бластомерів.
Вивчення індукційних взаємодій у різних представників типу хордових показало, що області і терміни компетенції неоднакові. Так, у асцидій на стадії 8 бластомерів, коли вже всі основні зачатки зумовлені, проводили деякі переміщення бластомерів. Матеріал хордомезодерми і основна частина нейрального матеріалу у них локалізовані в задньому вегетативному бластомер. Невелика частина нейрального матеріалу, що формує головний ганглій, знаходиться в задньому анімальному бластомер, розташованому над заднім вегетативним (рис. 8.10).
1 - епідерміс, 2 - нервова пластинка, 3 - хорда, 4 - ентодерми, 5 - соміти, 6 мезенхима
ДТШ перевірки індукційних взаємодій між ними анімальний ярус бластомеров повертали на 180 ° так, щоб задній анімальний бластомер втрачав контакт із заднім вегетативним. Головний ганглій не розвинувся ніде. Це означає, що для розвитку головного ганглія необхідно індукційне вплив на задній анімальний бластомер з боку заднього вегетативного. Крім того, очевидно, що задній анімальний бластомер не володіє автономністю розвитку, але тільки він компетентний до сприйняття впливу з боку заднього вегетативного бластомера, що містить хордомезодермальний зачаток.
У всіх інших класах хордових індукційні взаємодії між хордомезодермальний і нейтральні зачатками подібними до таких у амфібій. Вважають, що в ході еволюції хордових відбулися розширення областей іудліненіе терміну компетенції. Це розцінюють як ознака істотного еволюційного прогресу.
Індукційні численні і різноманітні. Крім первинної індукції з боку спинний губи бластопора описані індукційні впливу на більш пізніх, ніж гаструля- ція, етапах розвитку. Всі вони є вторинними і третинними,
бувши каскадні взаємодії, типові для диф- ференціровкі, тому що індукція багатьох структур залежить від попередніх індукційних подій. прикладом вторинної індукції може служити дію очного келиха (випинання переднього мозку) на прилеглий покривний епітелій, під впливом чого епітелій впячивается, а потім отшнуровивается кришталикових пузирек- зачаток очного кришталика (рис. 8.11). Розташований над кришталиком покривний епітелій теж відчуває складні зміни, втрачає пігмент і стає рогівкового епітелієм. Це приклад третинної індукції. Таким чином виходить, що очної келих виникає тільки після розвитку передньої частини головного мозку, кришталик - після формування келиха, а рогівка - після утворення кришталика.
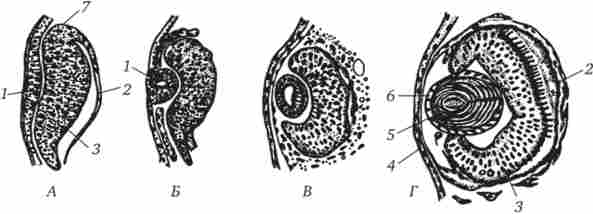
Мал. 8.11. Розвиток (А - Г) очі у хвостатої амфібії.
1 - кришталика плакода, 2 - пігментний епітелій, 3 - сітківка, 4 - рогівка, 5 - кришталикові волокна, 6 - кришталикових епітелій, 7 очної келих
Разом з тим індукція носить не тільки каскадний, а й переплітається характер, т. е. в індукції тієї чи іншої структури може брати участь не одна, а кілька тканин. У свою чергу, така структура може служити індуктором для кількох інших тканин. Наприклад, очної келих служить головним, але не єдиним індуктором кришталика. Морфогенез завжди супроводжується значними переміщеннями тканин один щодо одного. Так, презумптівного кришталик, т. Е. Епідерміс, з якого в подальшому повинен розвинутися кришталик, під час гаструляції лежить над ентодермою майбутньої глотки, яка служить першим індуктором кришталика. Потім під цим епідермісом виявляється серцева мезодерма, яка теж діє як індуктор. І тільки пізніше, під час нейруляции на передньому кінці нервової трубки, випинаються очні міхури, що утворюють очної келих і сітківку, що є головним індуктором кришталика (рис. 8.12).
Видаляючи ту чи іншу з індукують тканин, визначили ступінь участі кожної з них в індукції кришталика. Виявилося, що при видаленні сітківки очного келиха у 42% зародків амфібій
Мал. 8.12. Послідовні індукційні взаємодії, необхідні для утворення кришталика у зародка амфібії.
все ж формувалися кришталики і, отже, ентодерми і мезодерма в сумі володіють майже таким же индуцирующим дією, як і сітківка очного келиха.
I - ранній зародок, II - гаструла, III - нейрули, IV - стадія хвостовій нирки, V-личинка, VI - доросла особина; 1 - плакода, 2 - пухирець, 3 - волокна; а - ентодерми, б - серцева мезодерма, в - сітківка
Вважають, що численність індукують тканин може мати вирішальне значення для точного встановлення місця формування органу. Крім того, мережі індукції можуть відігравати важливу роль в каналізації розвитку, забезпечуючи нормальний перебіг органогенезу, навіть якщо один з компонентів индуцирующей системи не зуміє зробити сигнал потрібної сили.
Найчастіше прилеглі ділянки зародка надають взаємний вплив один на одного. Демонстративним прикладом є взаємодії в зародку кінцівки. Кінцівка розвивається зі скупчення клітин, що походять з бічної мезодерми, і покривають їх клітин ектодерми (рис. 8.13). Розвиток кінцівки починається з активації клітин бічної мезодерми в безпосередній близькості від сомітов, які, можливо, і надають індукують імпульси на мезодерму в області майбутньої кінцівки. Активовані мезо- дермальниє клітини зачатків кінцівки впливають на покриває їх ектодерму, в результаті чого вона потовщується. Утворене
Мал. 8.13. Вплив видалення апикального ектодермального гребеня на розвиток нирки крила. А - схема розташування нирки крила; Б - нирка крила; В - недорозвинення скелета (пунктир) після видалення апикального гребеня: потовщення епідермісу на його верхівці називають апікальним ектодермальних гребенем. Останній стимулює зростання нирки кінцівки (при видаленні його зростання нирки кінцівки припиняється). Мезодерма ж підтримує гребінь в активному стані і визначає форму кінцівки. Наприклад, мезодерма з нирки крила при з'єднанні з ектодермою нирки ноги утворює крило, вкрите пір'ям, або мезодерма з нирки кінцівки качиного зародка з ектодермою курячого призводить до розвитку перетинчастої кінцівки.
розрізняють гетерономний і гомономной види індукції. До гетерономной відносять випадки, подібні описаному, при яких один шматочок зародка індукує інший орган (хордомезодерми індукує поява нервової трубки і всього зародка в цілому). гомономная індукція полягає в тому, що індуктор спонукає навколишній матеріал до розвитку в тому ж напрямку, що і він сам. Наприклад, область нефротома, пересаджена іншому зародку, сприяє розвитку навколишнього матеріалу в сторону формування головного нирки, а поповнення в культуру фібробластів серця маленького шматочка хряща тягне за собою процес утворення хряща.
Щоб сприйняти дію індуктора, компетентна тканина повинна володіти хоча б мінімальної організацією. Поодинокі клітини не сприймають дію індуктора, а чим більше клітин в реагує тканини, тим активніше її реакція. Для надання індукуючогоефекту іноді достатньо лише однієї клітини індуктора.
Індукційні взаємодії можуть проявлятися в культурі тканини in vitro, але по-справжньому повноцінними вони бувають тільки в структурі цілісного організму.
Таким чином, індукційні виявлені на самих різних етапах розвитку багатьох хребетних. В акті індукції слід розрізняти два компоненти: індуктор і реагує область. Викладені вище положення коротко узагальнені на схемі 8.4.
Схема 8.4. закономірності індукції

В даний час інтенсивно ведуться роботи по вивченню молекулярних і клітинних механізмів індукції. У теоретичному сенсі явище ембріональної індукції допомагає по-новому оцінити взаємовідношення таких процесів, як залежна диференціювання ровка і детермінація, а також цітодіфференціровка і морфогенез. Поняття детермінації і морфогенезу більш детально розглядаються в розд. 8.3.