| Головна |
| «« | ЗМІСТ | »» |
|---|
Очевидно, що генетичний контроль розвитку існує, бо як тоді зрозуміти, чому з яйця крокодила розвивається крокодил, а з яйця людини - людина. Яким чином гени визначають процес розвитку? Це центральний і дуже складне питання, до якого вчені починають підходити, але для всеосяжного і переконливої відповіді на нього даних явно недостатньо. Головним прийомом вчених, які вивчають генетику індивідуального розвитку, є використання мутацій. Виявивши мутації, які змінюють онтогенез, дослідник проводить порівняння фенотипів мутантних особин з нормальними. Це допомагає зрозуміти, як даний ген впливає на нормальний розвиток. За допомогою численних складних і дотепних методів намагаються визначити час і місце дії гена. В даний час в біології розвитку використовується ряд новітніх методик, серед яких, наприклад, техніки knock out (Делеції гена) або knock down (Селективного придушення експресії гена за допомогою мРНК-антагоніста), метод FISH (застосування мічених фрагментів ДНК для виявлення певних молекул мРНК і їх розподілу в клітці і зародку). Вони дозволяють також визначити наявність взаємодії між генами і їх характер в ході ембріонального розвитку або на постембріональному етапі.
Аналіз генетичного контролю ускладнюється декількома моментами. Перш за все тим, що роль генів неоднакова. Частина геному складається з генів, що визначають так звані життєво важливі функції і відповідають, наприклад, за синтез тРНК або ДНК-полімерази, без яких неможливе функціонування жодної клітини. Ці гени названі «house keeping», або генами «домашнього господарства». Інша частина генів безпосередньо бере участь в детермінації, диференціювання і морфогенезі, т. Е. Функція їх, мабуть, більш специфічна, ключова.
Для аналізу генетичного контролю необхідно, крім того, знати місце первинного дії даного гена, т. Е. Слід розрізняти випадки відносної, або залежною, плейотропії від прямий, або істинної, плейотропії. У разі відносної плейотропії, як, наприклад, при серповидно-клітинної анемії, існує одне первинне місце дії мутантного гена - гемоглобін в еритроцитах, а всі інші спостережувані при ній симптоми, такі як порушення розумової та фізичної діяльності, серцева недостатність, місцеві порушення кровообігу, збільшення і фіброз селезінки і багато інших, виникають як наслідок аномального гемоглобіну. При прямий плейотропії все різноманітні дефекти, що виникають в різних тканинах або органах, викликаються безпосереднім впливом одного і того ж гена саме в цих різних місцях.
Нарешті, слід розрізняти ще два способи дії мутацій на фенотип, що викликають дизруптивного або гомеозісние зміни. У першому випадку, і це буває найчастіше, мутації призводять до порушення нормального розвитку, відсутності або аномальному будові органів. В інших випадках відхилення від норми полягає в тому, що під дією мутацій типовий орган заміщається
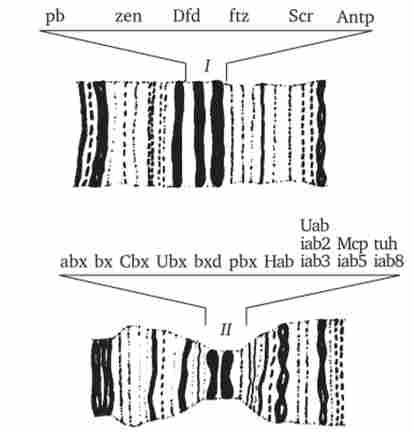
Мал. 8.14. Ділянки політенних хромосом дрозофіли із зазначенням розташування комплексів ANT-C (I) І ВХ-С (II) і генів, що входять в ці комплекси
гомологічним або зовсім іншим, але з нормальною будовою. Це особливий клас мутацій, описаний у комах і отримав назву гомеозісних мутацій.
Прикладом гомеозісних мутацій є мутації в ВХ-С і ANT-C-комплексах генів у дрозофіли (рис. 8.14). ВХ-С (Bithorax Complex) і ANT-C (Antennapedia Complex) - це два набори генів, що представляють собою два кластери тісно зчеплених між собою генів, що знаходяться в одному плечі 3-ї хромосоми (рис. 8.15). До групи ВХ-С входить не менше 10 генів, які відповідають за індивідуальність
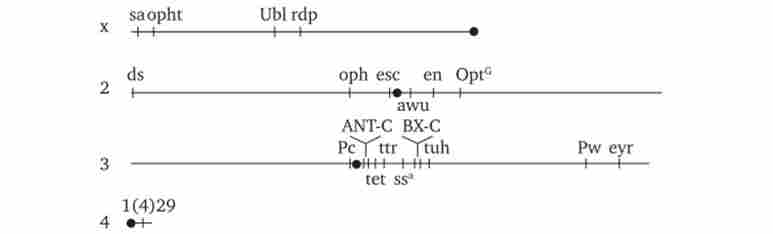
Мал. 8.15. Розташування гомеозісних комплексів мутантних генів ANT-C і ВХ-С в геномі дрозофіли.
X, 2, 3, 4 - Ікс-, друга, третя і четверта хромосоми; чорною крапкою позначені центромери, латинськими буквами - деякі гени
сегментів тіла, розташованих позаду середньогрудного сегмента. Чим більше дистальних локусів ВХ-С піддається делеции, тим більше черевних сегментів перетворюється в грудні. При делеции всього комплексу ВХ-С все сегменти від заднегрудного до 8-го черевного перетворюються в среднегрудь. До групи ANT-C входить не менше шести генів. Посилення в результаті мутації функції цієї групи викликає перетворення антени в ногу, як повинно бути в грудних сегментах.
Однак мутації по гомеозісним генам не змінюють у дрозофіли кількість або полярність сегментів. Звідси випливає, що гомео- зісние гени відповідають за якісну специфікацію сегментів тіла. Іншими словами, ці гени визначають, який сегмент (голови, грудей або черевця) і з якими їх структурами повинен сформуватися (рис. 8.16).
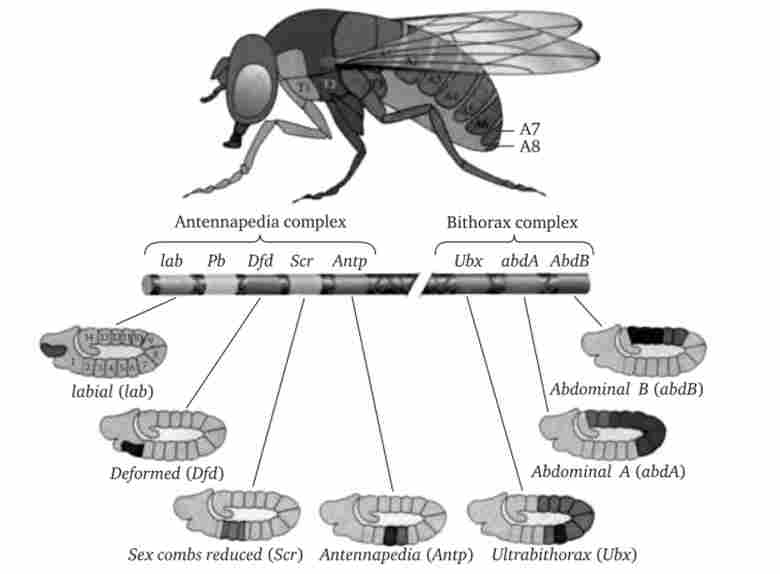
Мал. 8.16. Гомеозисні гени дрозофіли
Великі генетичні дослідження гомеозісних мутацій у дрозофіли дозволили ці комплекси генів визначити як селекторні гени. Виявилося, що продукти гомеозісних генів - це білки - фактори транскрипції, що регулюють транскрипцію генів початкових ланок генетичних формообразовательних програм. Продукти цієї групи генів далі впливають на експресію нижчестоящих генів-мішеней. Останні беруть участь в реалізації генетичної програми освіти конкретної структури або органи.
Слід зазначити, що гени, подібні до гомеозіснимі генами дрозофіли, знайдені у всіх інших тварин: від безхребетних (медузи) до хребетних, включаючи людину. Виявилося, що число кластерів гомеозісних генів в ході еволюції змінювалося (рис. 8.17).
Білки - продукти гомеозісних генів, через мутацій останніх у різних тварин змінили функції. Деякі гомеозисні гени в ході еволюції втрачені, інші - навпаки, дупліціроваться. Результатом мутацій цих генів у дрозофіли є втрата будь-яких сегментів тіла. При дослідженні таких мутацій були виявлені групи генів, названі генами сегментації. Продукти генів сегментації (як і продукти гомеозісних генів) транскрипційні фактори, послідовно контролюючі освіту сегментів тіла дрозофіли. Мутації генів групи gap призводять до втрати кількох прилеглих один до одного сегментів. Мутації в наступній за ними групі генів pair-rule - до втрати кожного другого сегмента. Нарешті, мутації генів сегментарной полярності - до заміни певних частин сегментів тіла структурами, що представляють дзеркальні відображення прилеглих половин сегментів.

Мал. 8.17. Зміна числа кластерів гомеозісних генів в еволюції у деяких груп тварин
Існують мутації, які вказують на існування у багатьох видів тварин так званих генів з материнським ефектом. Особливість цих генів полягає в тому, що материнський геном
під час овогенеза продукує ферменти, необхідні для метаболізму раннього зародка, а також передає інформацію, що стосується розташування і організації структур зародка, т. е. впливає на морфогенез. Тому самка, гомозиготна за рецесивним мутантного аллели і продукує аномальні яйця, навіть при схрещуванні з нормальним самцем дає нежиттєздатне потомство. Сама ж вона розвивалася цілком нормально, оскільки її мати в цьому випадку могла бути тільки гетерозиготною і в її яйцях були всі чинники, необхідні для раннього розвитку. Цікаво, що якщо в дефектні яйця рецессивной самки ввести цитоплазму від нормальних яєць, то зародки будуть врятовані. Фактори, детермініруемие генами з материнським ефектом, зазвичай роблять свій вплив на зародок до періоду гаструляції. Наприклад, це гени bicoid і banos, які беруть участь у формуванні передньо-задньої осі майбутнього зародка. Починаючи з гаструляції, все більшу роль відіграє інформація самого зародка. Наступність вищеперелічених генів в розвитку можна уявити на сьогоднішній день у вигляді спрощеної ієрархічної системи.
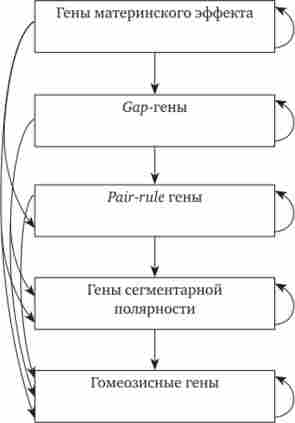
Мал. 8.18. Взаємодія генів, що контролюють сегментацію тіла в ранньому розвитку дрозофіли
Відомі й інші мутації, які впливають на ранній розвиток, але не пов'язані з материнським ефектом. До них відносяться, наприклад, мутації рибосомних генів. У шпорцевой жаби (Xenopus laevis) Мутанти були позбавлені частини або всіх рибосомних генів. Цитологічних це проявлялося в повній або частковій відсутності ядерця. У гомозиготному стані мутантних алелів зародки стають повністю нежиттєздатними на стадії вилуплення, так як у них не утворюються нові рибосоми, а ті, що були запасені в яйці, вже повністю використані.
У мишей також відомий цілий ряд рецесивних мутацій складного локусу Т 17-ої хромосоми, які зачіпають ранній розвиток. Локус Т представлений безліччю (117) алелей, які охоплюють знаком t з додатковими індексами: t1, t2, t3 і т. д. Близько 30% t-генів у гомозиготному стані викликає загибель зародків, частина алелей є напівлетальними. Весь цей ряд рецесивних алелей t розпадається на вісім груп, які можуть бути комплементарні один одному і в гетерозиготному стані не приводити до загибелі зародка.
Відомі також і п'ять домінантних мутацій Т-локусу. Кожна з восьми груп обумовлює різного роду дефекти. Один з алелей зупиняє перетворення морули в бластоцисту, що складається з трофобласта і ембріобласта. Такі морули гинуть. Інша мутація призводить до того, що розвинувся трофобласт не вступає в контакт зі стінкою матки і зародок теж гине. Треті мутантні зародки не утворюють внезародишевой ектодерми, у четвертих - гинуть клітини зародкової ектодерми, у п'ятих - клітини зародкової ектодерми не здатні мігрувати в області первинної смужки і утворювати мезодерму, у шостих - вже утворилися структури нервової системи дегенерируют і т. Д. Первинне порушення, лежить в основі всіх цих ефектів, всього лише одного локусу поки не з'ясовано. Однак очевидно, що локус Т грає першорядну роль в морфогенезі ектодерми мишачого зародка і організму в цілому.
Відомі мутації з більш специфічною дією. У аксолотля мутація з (cardial lethal), що успадковується як простий аутосомно-рецес- вибухобезпечний ознака, викликає порушення розвитку серця і його нездатність до скорочень. Мутантні особини роздуті переповнює їх рідиною, а травна система і зябра у них недорозвинені. Встановлено, що у хребетних серце розвивається в результаті індукційного впливу з боку головного ділянки ентодерми. Досліди з пересадки серцевої мезодерми показали, що мутантні зародки не можуть забезпечити розвиток серця з підсадженої нормальної серцевої мезодерми. Це вказує на відсутність індукційного активності головного ділянки ентодерми у мутанта з генотипом сс.
Нерозривний зв'язок генів з морфогенезом окремих органів і систем органів можна спостерігати також на прикладі мутації Tfm (Testicular feminization locus) у ссавців. У людини, мишей і щурів цей ген успадковується як зчеплена з підлогою. Самки, гетерозиготні по мутантного гену, т. Е. X7fmX, по суті, нормальні, але половина їх генотипично чоловічих нащадків мають жіночий фенотип і стерильні. В основі цього дефекту лежить відсутність специфічного білка, службовця рецептором тестостерону. У особин XTfmY цей рецептор не синтезується (див. Також розд. 3.6.5.2 і 7.1.2.1). самки XTfmXTfm нормальні в усіх відношеннях і фертильності.
Це свідчить про те, що нормальне статеве розвиток самки може проходити і без продукту цього гена.
Органогенез - період, коли дія мутацій проявляється у великій мірі. Розвиток кожного органу і тим більше системи органів контролюється сукупним координованим дією сотень генів. У людини відомі понад 120 форм спадкової глухоти, які виникають в результаті експресії мутантних генів, що відповідають за формування слухового аналізатора. У людини описано також близько 250 спадкових уражень очей, близько 150 спадкових аномалій розвитку скелета, не менше 18 генів, що відповідають за нормальну диференціацію статі. Про значення генетичного контролю онтогенезу говорять численні хвороби, пов'язані з геномних і хромосомними мутаціями.
В цілому генетичний контроль онтогенезу очевидний, проте в процесі розвитку зародок і його частини мають здатність до саморозвитку, регульованим самої цілісної системою, що розвивається і не запрограмованому в генотипі зиготи.