| Головна |
| «« | ЗМІСТ | »» |
|---|
Диференціація - це процес, в результаті якого клітина стає спеціалізованої, т. е. набуває хімічні, морфологічні та функціональні особливості. В самому вузькому сенсі - це зміни, що відбуваються в клітині протягом одного, нерідко термінального, клітинного циклу, коли починається синтез головних, специфічних для даного клітинного типу, функціональних білків (схема 8.1). Прикладом може служити диференціювання клітин епідермісу шкіри людини, при якій в клітинах, які прямують з базального в шипуватий і потім послідовно в інші, більш поверхневі шари, відбувається накопичення кера- тогіаліна, що перетворюється в клітинах блискучого шару в елеідін, а потім в роговому шарі - в кератин. При цьому змінюються форма клітин, будова клітинних мембран і набір органоїдів. Насправді диференціюється не одна клітина, а група подібних клітин. Прикладів можна навести безліч, так як в організмі людини налічують близько 220 різних типів клітин. Фібробласти синтезують колаген, міобласти - міозин, клітини епітелію травного тракту - пепсин і трипсин.
У більш широкому сенсі під дифференцировкой розуміють поступове (протягом декількох клітинних циклів) виникнення все великих відмінностей і напрямків спеціалізації між клітинами, які відбулися з більш-менш однорідних клітин одного вихідного зачатка. Цей процес неодмінно супроводжують морфогенетичні перетворення, т. Е. Виникнення і подальший розвиток зачатків певних органів в дефінітивного органи. Перші хімічні та морфогенетичні відмінності між клітинами, що обумовлюються самим ходом ембріогенезу, виявляються в період гаструляції.
Процес, в результаті якого окремі тканини в ході дифферен- цировки набувають характерного для них вид, називають гістогенезом. Диференціація клітин, гістогенез і органогенез відбуваються в сукупності, причому в певних ділянках зародка і в певний час. Це дуже важливо, тому що вказує на координованість і інтегрованість ембріонального розвитку.
Необхідно зрозуміти, яким чином клітини, що володіють найчастіше однаковими кариотипом і генотипом, диференціюються і беруть участь в гисто- і органогенезу в необхідних місцях і в певні терміни відповідно цілісного «образу» даного виду організмів. Обережність при висуненні положення про те, що
Глава 8. Закономірності індивідуального розвитку організмів Схема 8.1. Диференціація мезодерми
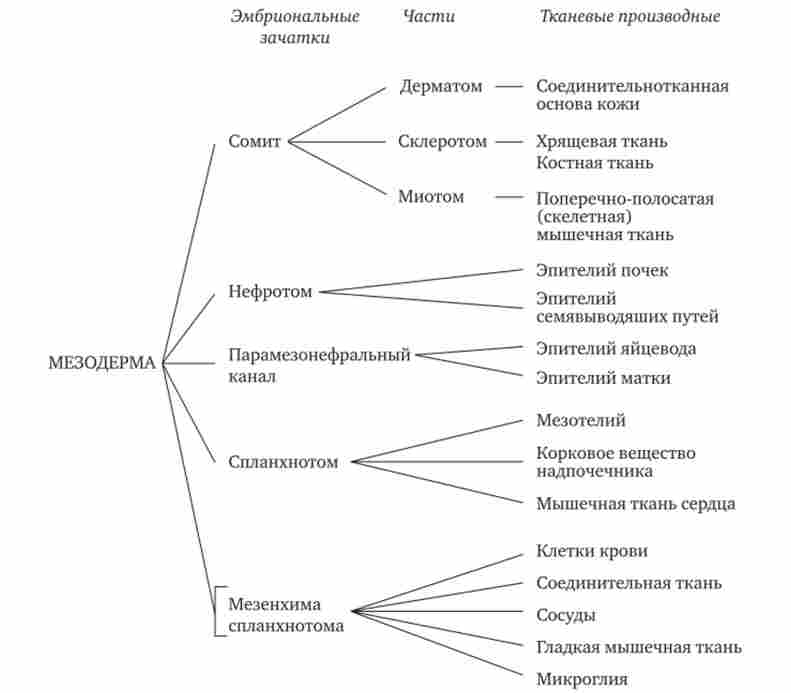
спадковий матеріал усіх соматичних клітин абсолютно ідентичний, відображає об'єктивну реальність і історичну неоднозначність в трактуванні причин клітинної диференціювання. Розвиток уявлень про механізми цітодіфференціровкі зображено на схемі 8.2.
В. Вейсман висунув гіпотезу (кінець XIX ст.) Про те, що тільки лінія статевих клітин несе в собі і передає нащадкам всю інформацію свого генома. Соматичні клітини, на його думку, можуть відрізнятися від зиготи і один від одного кількістю спадкового матеріалу і тому диференціюватися в різних напрямках.
Пізніше були виявлені приклади зміни кількості спадкового матеріалу в соматичних клітинах як на геномном, так і на хромосомному і генному рівнях. Описані випадки елімінації цілих хромосом у циклопа, комара і у одного з представників сумчастих. У останніх із соматичних клітин самки елімінується Х-хромосома, а з клітин самця - Y-хромосома. В результаті соматичні клітини у них містять лише по одній Х-хромосомі, а в лінії статевих клітин зберігаються нормальні каріотипи: XX або XY.
Схема 8.2. Розвиток уявлень про механізми цітодіфференціровкі

У політенних хромосомах слинних залоз двокрилих ДНК може синтезуватися несинхронно, наприклад при політенізаціі гетерохроматіновие ділянки реплицируются менше число раз, ніж еухроматіновие. Сам процес політенізаціі, навпаки, призводить до значного збільшення кількості ДНК в диференційованих клітинах в порівнянні з родоначальних клітинами.
Такий механізм реплікації ДНК, як ампліфікація, також призводить до багаторазового збільшення кількості деяких генів в одних клітинах в порівнянні з іншими. У овогенезі багаторазово збільшується число рибосомних генів, можуть ампліфіціро- тися і деякі інші гени. Є дані про те, що в деяких клітинах в процесі диференціювання відбувається перебудова генів, наприклад іммуноглобулінових генів в лімфоцитах.
Однак в даний час загальновизнаною є точка зору, провідна початок від Т. Моргана, який, спираючись на хромосомну теорію спадковості, припустив, що діффе- ренціровка клітин в процесі онтогенезу є результатом послідовних реципрокних (взаємних) впливів цитоплазми і мінливих продуктів активності ядерних генів. Таким чином, вперше прозвучала ідея про диференціальної експресії генів
як основний механізм цітодіфференціровкі. В даний час зібрано багато доказів того, що в більшості випадків соматичні клітини організмів несуть повний диплоїдний набір хромосом, а генетичні потенції ядер соматичних клітин можуть зберігатися, т. Е. Гени втрачають потенційної функціональної активності.
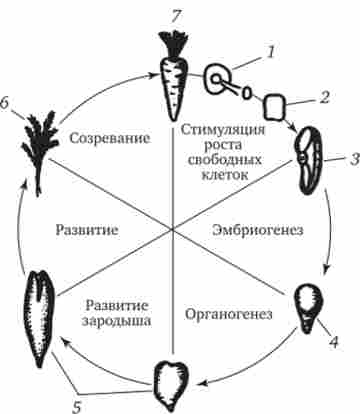
Мал. 8.6. Досвід, який показує збереження функціональних властивостей спадкового матеріалу в соматичної диференційованої клітці моркви.
1 - зріз кореня в живильному середовищі, 2 - профілюючі клітини в культурі, 3 - клітина, ізольована з культури, 4 - ранній зародок, 5 - пізніший зародок, 6 - молоде рослина, 7-доросла рослина
Збереження повного хромосомного набору, що розвивається забезпечується, насамперед, механізмом мітозу. Про збереження генетичних потенцій ядер соматичних клітин можна судити за результатами дослідів, проведених над рослинами і тваринами. Минулий тривалий шлях диференціювання соматична клітина моркви здатна розвиватися в повноцінний організм (рис. 8.6). У тварин окремі соматичні клітини після стадії бластули, як правило, не здатні розвиватися в цілий нормальний організм, але їх ядра, будучи пересаджені в цитоплазму овоциту або яйцеклітини, починають вести себе відповідно тієї цитоплазмі, в якій вони опинилися.
Досліди по пересадці ядер соматичних клітин в яйцеклітину вперше були успішно здійснені в 1950-х рр. в США, а в 1960- 1970-х рр. отримали широку популярність досліди англійського вченого Дж. Гердон. Використовуючи африканську шпорцевую жабу Xenopus laevis, він в невеликому відсотку випадків отримав розвиток дорослої жаби з енуклійованої яйцеклітини, в яку пересаджував ядро з епітеліальної клітини шкіри жаби або кишечника пуголовка, т. е. з диференційованої клітини (див. рис. 5.3). Енуклеація яйцеклітини проводили великими дозами ультрафіолетового опромінення, що призводило до інактивації її ядра. Для доказу того, що в розвитку зародка бере участь пересаженное ядро соматичної клітини, застосували генетичне маркування. Яйцеклітину брали з лінії жаб з двома ядерця в ядрі, а ядро клітини донора - з лінії, що має в ядрах тільки одне ядерце внаслідок гетерозиготности по делеции ядерцевого організатора. Всі ядра в клітинах особини, отриманої в результаті трансплантації ядра, мали тільки одне ядерце.
Разом з тим досліди Гердон виявили багато інших найважливіших закономірності. По-перше, вони ще раз підтвердили припущення Т. Моргана про вирішальне значення взаємодії цитоплазми і ядра в життєдіяльності клітин і розвитку організму. По-друге, в численних експериментах було показано, що чим старше стадія зародка-донора, з клітин якого брали ядро для пересадки, тим в меншій відсотку випадків розвиток виявлялося повністю завершеним, т. Е. Досягало стадій пуголовка, а потім жаби.
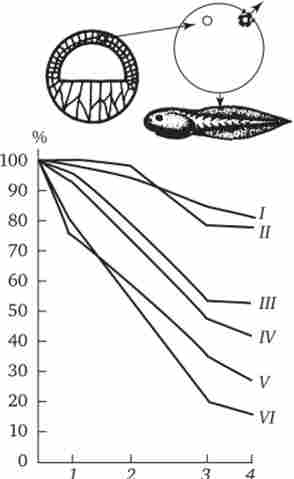
Мал. 8.7. Залежність успіху пересадки ядер з диференційованої клітини в яйцеклітину від віку донора (I - VI) ядра.
про - | -1-1-1
12 3 4
Стадія розвитку, що досягається кліткою-реципієнтом ядра
У більшості випадків розвиток зупинялося на більш ранніх стадіях. Залежність результатів пересадки від стадії зародка-донора ядер представлена на рис. 8.7. Аналіз зародків, що зупиняються в розвитку після пересадки ядра, показав безліч хромосомних аномалій в їх ядрах. Іншою причиною зупинки розвитку вважають нездатність ядер диференційованих клітин до відновлення синхронної реплікації ДНК.
Головний висновок, який випливає з цього досвіду, полягає в тому, що спадковий матеріал соматичних клітин здатний зберігатися повноцінним не тільки в кількісному, але і в функціональному відношенні, цітодіффе- ренціровка не є наслідком недостатності спадкового матеріалу.
Експерименти з клонування рослин і тварин - доказ повноцінності матеріалу соматичної клітини. Вчені не виключають можливості відтворення подібним вівці Доллі чином, т. Е. Шляхом пересадки ядер, генетичних двійників людини. Слід, однак, віддавати собі звіт, що клонування людини крім науково-технологічного має також етичний і психологічний аспекти.
Гіпотеза диференціальної експресії генів в ознака приймається в даний час в якості основного механізму цітодіф- ференціровкі.
Рівні регуляції диференціальної експресії генів відповідають етапам реалізації інформації в напрямку ген -> поліпептид - е ознака і включають не тільки внутрішньоклітинні процеси, але тканинні і організменние.
Експресія гена в ознаку - це складний етапний процес, який можна вивчати різними методами: електронної та світловій мікроскопії, біохімічно і іншими. На схемі 8.3 наведені основні етапи експресії генів і методи, за допомогою яких їх можна вивчати.
схема 8.3
|
Етапи експресії генів |
Методи їх вивчення |
|
активність генів |
Візуальне спостереження будови відповідних ділянок хромосоми (електронна і світлова мікроскопія) |
|
Транскрипція, первинний РНК-транскрипт (ядерні РНК) |
Метод двовимірного гельелектрофореза |
|
мРНК цитоплазми |
біохімічний метод |
|
Трансляція (білки - продукти генної активності) |
біохімічний метод |
|
морфологічна диференціювання |
Цитологічний метод, цитохимичний метод |
|
Будова і життєздатність зародка |
Гибридологічний і порівняно ембріологічний методи |
Візуальне спостереження в електронний мікроскоп проведено відносно тільки окремих генів - Хвороби, генів хромосом типу лампових щіток і деяких інших (див. Рис. 3.66). На електронограммах чітко видно, що одні гени транскрибуються активніше за інших. Добре помітні і неактивні гени.
Особливе місце займає вивчення політенних хромосом. політенія - це гігантські хромосоми, виявлені в інтерфазних клітинах деяких тканин у мух і інших двокрилих. Такі хромосоми є у них в клітинах слинних залоз, мальпігієвих судин і середньої кишки. Вони містять сотні ниток ДНК, які редуплікованих, але не піддалися розбіжності. При фарбуванні в них виявляються чітко виражені поперечні смуги або диски (див. Рис. 3.56). Багато окремі смуги відповідають місцю розташування окремих генів. Обмежене число певних смуг в деяких диференційованих клітинах утворює здуття, або пуфи, які виступають за межі хромосоми. Ці роздуті ділянки знаходяться там, де гени найбільш активні щодо
транскрипції. Було показано, що клітини різного типу містять різні пуфи (див. Рис. 3.65). Зміни в клітинах, що відбуваються в ході розвитку, корелюють зі змінами в характері пуфів і синтезом певного білка. Інших прикладів візуального спостереження генної активності поки немає.
Всі інші етапи експресії генів є результатом складних видозмін продуктів первинної генної активності. Під складними змінами на увазі посттранскрипційна перетворення РНК, трансляцію і Посттрансляційні процеси.
Є дані по вивченню кількості і якості РНК в ядрі і цитоплазмі клітин організмів, що знаходяться на різних стадіях ембріонального розвитку, а також в клітинах різних типів у дорослих особин. Виявлено, що складність і число різних видів ядерної РНК в 5-10 разів вище, ніж мРНК. Ядерні РНК, які представляють собою первинні продукти транскрипції, завжди довше, ніж мРНК. Крім того, ядерна РНК, вивчена на морському еже, за кількістю і якісному розмаїттям ідентична на різних стадіях розвитку особини, а мРНК цитоплазми відрізняється в клітинах різних тканин. Це спостереження наводить на думку про те, що посттранскрипційна механізми впливають на диференціальну експресію генів.
Приклади посттранскрипційна регуляції експресії генів на рівні процесингу відомі. Мембранно-пов'язана форма імуноглобуліну IgM у мишей відрізняється від розчинної форми додаткової амінокислотною послідовністю, що дозволяє мембранно-зв'язаній формі «заякорюють» в клітинній мембрані. Обидва білка кодуються одним локусом, але процесинг первинного транскрипту протікає по-різному. Пептидний гормон кальцитонін у щурів представлений двома різними білками, детермінованими одним геном. У них однакові перші 78 амінокислот (при загальній довжині 128 амінокислот), а відмінності обумовлені процессингом, т. Е. Знову спостерігається диференціальна експресія одного і того ж гена в різних тканинах. Є й інші приклади. ймовірно, альтернативний процесинг первинних транскріп- тов грає дуже важливу роль в диференціюванні, однак залишається неясним його механізм.
Велика частина мРНК цитоплазми однакова за якісним складом в клітинах, що відносяться до різних стадіях онтогенезу; мРНК необхідні для забезпечення життєдіяльності клітин і детермінуються генами «домашнього господарства», представленими в геномі у вигляді декількох нуклеотидних послідовностей із середньою частотою повторюваності. Продуктами їх активності є білки, необхідні для збірки клітинних мембран, різних субклітинних структур і т. Д. Кількість цих мРНК складає приблизно 9/10 від всіх мРНК цитоплазми. Решта мРНК є необхідними для певних стадій розвитку, а також різних типів клітин.
При вивченні різноманітності мРНК в нирках, печінці та головному мозку мишей, в яйцеводах і печінки курей було виявлено близько 12 000 різних мРНК. Лише 10-15% були специфічні для будь-якої однієї тканини. вони зчитуються з унікальних нуклеотидних послідовностей тих структурних генів, дія яких специфічно в даному місці і в даний момент і які називаються генами «розкоші». Кількість їх відповідає приблизно 1000-2000 генів, відповідальних за диференціювання клітин.
Не всі гени, наявні в клітині, взагалі реалізуються до етапу освіти мРНК цитоплазми, але і ці утворилися мРНК не всі і не у всяких умовах реалізуються в поліпептиди і тим більше в складні ознаки. Відомо, що деякі мРНК блокуються на рівні трансляції, будучи в складі рібонуклеопротеінових частинок - інформосом, внаслідок чого відбувається затримка трансляції. Це має місце в овогенезі, в клітинах кришталика ока.
У ряді випадків остаточна диференціювання пов'язана з «добудовою» молекул ферментів або гормонів або четвертинної структури білка. Це вже Посттрансляційні події. Наприклад, фермент тирозинази з'являється у зародків амфібій ще в ранньому ембріогенезі, але переходить в активну форму лише після їх вилуплення.
Диференціація клітин не зводиться тільки до синтезу специфічних білків, тому стосовно багатоклітинні організми ця проблема невідривно від просторово-часових аспектів і, отже, від ще більш високих рівнів її регуляції, ніж рівні регуляцій біосинтезу білка на клітинному рівні. Диференціація завжди зачіпає групу клітин і відповідає завданням забезпечення цілісності багатоклітинного організму.