| Головна |
| «« | ЗМІСТ | »» |
|---|
Виходячи зі сказаного в попередньому параграфі, виникає питання: якщо в довгастому мозку і мосту нейрони рогів СМ перетворюються в ядра черепних нервів, то чому ж стає його проміжне ядро? Нагадаємо, що ця область є найбільш медіальної частиною сірої речовини СМ і найважливішим центром, який приймає рішення про запуск реакцій.
На це запитання можна дати таку відповідь: при переході від СМ до стовбурових структур проміжне ядро в цілому зберігає свою локалізацію, а також командні «обчислювальні» функції, і дає початок численним ретикулярним ядер (ретикулярної формації довгастого мозку, моста і середнього мозку).
Отже, в серединній частині стовбура мозку розташована ретикулярна формація (РФ) - скупчення нейронів різних розмірів і форм, розділених безліччю проходять в різних напрямках волокон, що нагадують мережу (лат. reticulum). Окремі ядра всередині РФ на анатомічному рівні визначаються з працею, оскільки зазвичай не розмежовані чіткими зонами білої речовини. В основі поділу РФ на ядра лежить, перш за все, функціональний принцип: за подібністю і розбіжності вирішуваних завдань, а також аферентів і ефферентов.
Важливим загальним властивістю більшості нейронів РФ є їх форма і характер організації зв'язків. Нейрони РФ є клітинами типу Гольджі I. Їх аксони часто мають дві гілки, перша з яких йде рострально, а друга - каудально. Отже, від ретикулярних нейронів можуть одночасно починатися висхідні і низхідні шляхи, що передають імпульси до багатьох структур ЦНС (рис. 6.6).
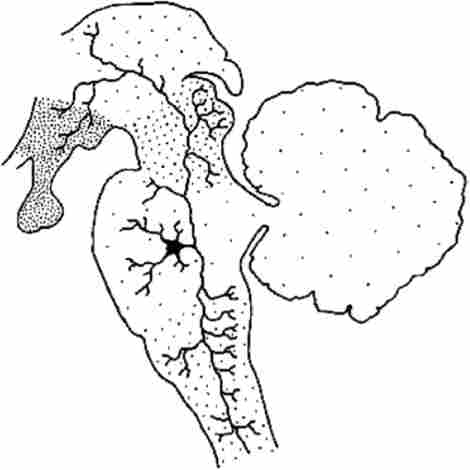
Мал. 6.6. Приклад розгалуження нейрона ретикулярної формації моста
Довгі розгалужені дендрити нейронів РФ орієнтовані переважно у фронтальній площині. З ними, як правило, утворюють синапси волокна, що несуть сигнали від різних сенсорних систем (слуховий, зорової, тактильної і т. Д.) І внутрішньомозкових областей. Іншими словами, для РФ надзвичайно характерна конвергенція (сходження) різноманітних інформаційних потоків.
У зв'язку з такими особливостями (реакцією на численні сигнали і формуванням ефферентов до великої кількості центрів) РФ в середині XX в. назвали неспецифічної. При цьому її протиставили специфічним системам, які приймають інформацію від вузького кола структур і передають її далі по конкретним «адресами».
В даний час відомо, що ретикулярні ядра, як і інші області ЦНС, цілком специфічні за своїми функціями. Разом з тим, функції ці дійсно вимагають одночасного обліку великої різноманітності сигналів, а також участі в реалізації запускаються реакцій безлічі мозкових структур (аж до кори великих півкуль). Сказане відноситься до діяльності ретикулярних ядер, що регулюють харчову поведінку і захисні реакції, до Судиноруховий і дихальним центрам. Але в максимальному ступені це справедливо для областей, які керують сном і неспанням.
Так, для центрів неспання (основний з них знаходиться в РФ моста) дуже важливо мати можливість реагування на все розмаїття сенсорних сигналів. В результаті ми прокидаємося від гучного звуку, яскравого світла, тактильного роздратування і т. П. Однак не менш важливо, щоб при зростанні рівня неспання активізувалася вся ЦНС - і цієї мети служать широко розгалужені колатералі нейронів ретикулярних ядер моста.
За структурно-функціональним критеріям РФ підрозділяється на три зони, медіанну, медіальну і латеральну. Медіанна зона лежить на середній лінії, медійна - всередині і в центрі стовбура, а латеральна - поблизу сенсорних ядер. У медіанної зоні присутні переважно дрібні нейрони, в медіальній зоні - великі і навіть гігантські нейрони, а в латеральної - нейрони середнього розміру. Медіанна зона тягнеться від довгастого мозку до каудальних (задніх) відділів середнього мозку (див. Рис. 6.2 і 6.5). Структури, що входять в цю зону, об'єднуються під загальною назвою ядер шва. В середньому мозку до ядер шва примикають ядра центрального сірої речовини, по багатьом особливостям будови і функцій подібні до ядрами РФ (див. Гл. 7). Для нейронів ядер шва характерно присутність в якості медіатора такого з'єднання, як серотонін.
Основний обсяг сенсорної аферентації РФ отримує від чутливих ядер черепних нервів і висхідних шляхів СМ (перш за все, спинно-ретикулярного). Крім того, на ретикулярних нейронах утворюють синапси колатералі низхідних трактів (кортико-спинального, рубро- спинального і ін.). Окремі області РФ вельми специфічно пов'язані з мозочком і управлінням рухами.
Ефферентов РФ утворюють висхідну і спадну системи волокон. Висхідні аксони йдуть в передній мозок - до медіальний ядер таламуса (проміжний мозок), корі великих півкуль. Спадні аксони направляються в СМ (ретикуло-спинальний тракт); волокна ретикулярних нейронів йдуть також до мозочка.
Численні зв'язку мають місце і всередині РФ між різними її областями, а також між ретикулярними ядрами та іншими стовбуровими структурами.
Таким чином, ретикулярна формація в значній мірі регулює роботу всієї ЦНС і забезпечує виконання найважливіших інтеграційних функцій. Дані функції досить численні і на сьогоднішній день ще не до кінця досліджені. Як вже було сказано, РФ грає ключову роль в управлінні рівнем загальної активності НС і забезпечує регулювання циклу «сон - неспання». При цьому різні ретикулярні ядра, переважно пов'язані зі сном або неспання, знаходяться у взаємно конкуруючих відносинах; враховують не тільки сенсорні сигнали із зовнішнього світу, а й інформацію від внутрішнього середовища організму (наприклад, про хімічний склад крові); реагують на добові та сезонні ритми.
Через шляху, котрі пов'язують РФ зі СМ і мозочком, вона бере участь в управлінні позою і локомоцией. Нагадаємо, що ретикуло-спинальний тракт впливає на роботу м'язів тулуба, плечового і тазового поясів. У риб він запускає швидкі вигини тіла при плаванні (за рахунок ритмічної імпульсної активності особливих гігантських нейронів).
В якості найважливішої функції РФ слід назвати регуляцію дихання. Усередині довгастого мозку і моста існують особливі центри вдиху і видиху, які підтримують і змінюють (частішають або урежают) дихальний ритм з урахуванням концентрації 02 і С02 в крові, ступеня розтягування легенів, рівня неспання, емоцій, команд з кори великих півкуль, завдяки чому можливий довільний контроль дихання і т. д. Імпульси від центрів вдиху і видиху направляються в СМ - до проміжного ядра і мотонейронам, розташованим в шийних і грудних сегментах спинного мозку.
Ще одна життєво важлива функція РФ - управління роботою серця і тонусом судин. Відповідні ретикулярні ядра об'єднують в судиноруховий центр довгастого мозку і моста. Тут важливо врахувати, що, на відміну від дихальних рухів, що запускаються ЦНС, серце б'ється самостійно (володіє автоматией). Судиноруховий центр за допомогою ВНС може лише «рекомендувати» йому змінити частоту і силу скорочень. При цьому, як і в випадку дихання, враховуються концентрація 02 і С02 в крові, емоції, рівень неспання. Дуже велике значення має інформація про ступінь розтягу основних судин нашого організму (аорти, порожнистих вен). Довільний контроль діяльності судинного центру (на відміну від дихального) практично відсутня.
Через РФ проходять рефлекторні дуги, що забезпечують реалізацію вроджених харчових і оборонних реакцій: смоктання, ковтання, слиновиділення, виділення шлункового соку, кашлю, чхання, блювання та ін. При цьому запускають сенсорні сигнали надходять переважно через чутливі ядра черепних нервів; виконання рефлексів забезпечують їх рухові і вегетативні ядра. Це ще раз показує, що, хоча стовбур (в першу чергу, довгастий мозок і міст) є складовою частиною ГМ, його будова і принципи функціонування в значній мірі ще ті ж, що і в разі СМ. Зокрема, основний інтеграційний комплекс ядер - РФ - відповідає проміжному ядру сірої речовини СМ.
Оскільки ростральна частина РФ розташовується в середньому мозку, ряд пов'язаних з нею питань додатково розглянуто в гл. 7. Робота дихального і судинного центрів буде детально описана в частинах підручника, присвячених відповідним системам організму людини (дихальної та серцево-судинної). Діяльність центрів сну і неспання також стане предметом окремого обговорення.