| Головна |
| «« | ЗМІСТ | »» |
|---|
У попередньому розділі представлена описова модель роботи скелетних м'язів. Наступний етап системного аналізу м'язи - дослідження структурних станів окремих елементів саркомерних містків.
Виділяють 6 основних станів саркомерного містка. Ці стани визначають за часом перебування в них системи. При цьому виділяються найбільш стабільні (довгостроково існуючі) стану. В основі роботи містка лежить реакція гідролізу аденозілтріфосфата - субстрату:
де АТФ, АДФ - аденозілтрі- і аденозілдіфосфат, Р / - фосфат (неорганічний фосфор), ДО "- енергія Гіббса реакції при фізіологічних умовах.
Щоб саркомерний місток працював, потрібно додати кальцій. Крім того, необхідні актин і міозин.
Гідроліз протікає при наявності трьох хімічних компонентів: кальцію, актину і міозину. Ферментом Е в цій реакції є міозин. Актин - аллостерічеський ефектор. Це означає, що за відсутності актину миозин діє дуже неефективно: повільно розкладає аденозілтріфосфат. І, в свою чергу, актин «не працює» без кальцію.
В результаті біохімічної реакції цих речовин виходять АДФ і Р ,.
АТФ - це субстрат ферменту Е. Субстрат надходить в активний центр (ключ в замок, див. рис. 8.13), в якому гідролізується. Енергія Гіббса даної реакції при стандартних біологічних умовах становить приблизно -30 кДж / моль.
Як уже зазначалося, саркомерний місток може перебувати в 6 різних станах.
Стан 1. На МІОЗИНУ немає ні субстрату АТФ, ні продуктів - АДФ і фосфату Р ,. Місток розімкнути, зачеплення-замикання немає.
Стан 2. Міозин (фермент Е) з'єднується з субстратом АТФ, замикання немає:
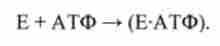
Стан 3. Субстрат АТФ розпадається, міозин з'єднаний з продуктами:
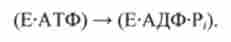
У станах 1-3 місток розімкнути.
Стан 4. До міозин не приєднався субстрат, ні продукт.
Стан 5. До міозин приєднаний субстрат АТФ.
Стан 6. З миозином пов'язаний АТФ і неорганічний фосфат Р "відбувається замикання.
У станах 4-6 місток замкнутий.
Результатом цих біохімічних реакцій є сукупність переходів між стабільними станами саркомерного містка (табл. 13.1).
Перехід зі стану 1 в стан 2.
Для цього необхідно, щоб миозин приєднав субстрат:
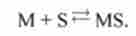
В результаті реакції утворюється міозин-субстратної комплекс MS. Ця реакція оборотна: можливий процес, що протікає в зворотному напрямку. Міозін- субстратної комплекс може розпадатися (про це нагадує подвійна стрілка в першому стовпці табл. 13.1). Тут константа швидкості прямої реакції (перехід I -> 2) позначена <Х|2, константа скорости обратной реакции (переход 2 ->) - <хг|.
Перехід зі стану 1 в стан 3.
Міозин приєднує продукт:
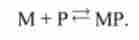 Таблиця 13.1
Таблиця 13.1
Переходи між станами саркомерного містка
|
перехід |
реакція |
Константи швидкості реакції |
Примітки |
|
1 2 |
M + S ^ MS |
«12» «21 |
|
|
1 3 |
М + Р ^ МР |
«13» «31 |
|
|
1 4 |
М + А ^ МА |
а, 4.041 |
|
|
2 3 |
MS ^ МР |
«23» «32 |
При додаванні інгібітора - холостий хід містка |
|
2<-»5 |
MS + Ai MSA |
«25» «52 |
|
|
3 6 |
МР + А "= * MPA |
«36» «63 |
|
|
4 5 |
MA + S ^ MSA |
«45» «54 |
|
|
4 «-> 6 |
МА + Р ^ MPA |
«46» «64 |
|
|
5<-*6 |
MSA ^ MPA |
«56» «65 |
Робочий хід містка: ніжка міозину згинається, міозин «замикається» на актине, субстрат розпадається |
Примітка: М - міозин, А - актин, S - субстрат АТФ, Р - продукт АДФ + Р ,.
Реакція М + Р »МР і зворотна реакція: розпад комплексу міозин - продукт на вільний міозин і продукт. Константа швидкості прямої реакції - сиз, константа швидкості зворотної реакції - аз).
Перехід зі стану 1 в стан 4.
Міозин реагуєте актином:
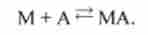
Місток замикається, але робота не відбувається. Константи швидкості прямої і зворотної реакцій - <Х|4, cui соответственно.
Перехід зі стану 2 в стан 3:
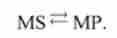
Перехід обумовлений розпадом субстрату на міозину. Реакція протікає без замикання, що відповідає так званому холостому ходу містка. Цей перехід спостерігається в тих випадках, коли відбувається розпад А IФ, проте м'яз при цьому не працює. Константи швидкості прямої і зворотної реакцій - АГЗ, азг-
Перехід зі стану 2 в стан 5.
Міозин захоплює субстрат і замикається на актин:
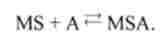
Відбувається замикання містка. Константи швидкості прямої і зворотної реакцій - <Х25, СХ52 .
Перехід зі стану 3 в стан 6.
Міозин з продуктами замикається на актин і назад:
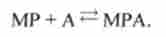
Константи швидкості прямої і зворотної реакцій - а.%, <х«з.
Перехід зі стану 4 в стан 5.
Міозин з'єднаний з актином, приєднує субстрат, утворюється гройной комплекс: міозин, субстрат, актин:
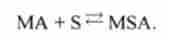
Починається робота містка. Можлива зворотна реакція: потрійний комплекс може розпадатися. Константи швидкості прямої і зворотної реакцій - ajs »CI54.
Перехід зі стану 4 в стан 6.
Міозин-актинового комплекс приєднує продукт, можлива зворотна реакція:
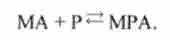
Константи швидкості прямої і зворотної реакцій - а ^.
Перехід зі стану 5 в стан 6.
У реакції беруть участь міозин, продукт, актин:
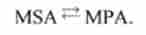
Константи швидкості прямої і зворотної реакцій - as *, (Хб5-
Ця остання, найважливіша реакція і є тим переходом, під час якого миозин замкнутий на актин.
На міозину є субстрат. Субстрат розпадається. Енергія ДG '= -30 кДж / моль цього розпаду виділяється. Відбувається робітничий рух саркомерного містка (рис. 13.7). Саме з переходів безлічі саркомерних містків складається робота м'язи.
Динаміку переходів в біохімічної системі з кінцевим набором стабільних станів наочно представляє кінетичний граф (Рис. 13.8).
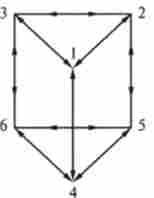
Мал. 13.8. Кінетичний граф саркомерного містка
Граф - сукупність точок (вершини) і що з'єднують їх ліній. Якщо лінії орієнтовані, то їх називають гілками; якщо лінії не орієнтовані, то їх називають ребрами. Кожній гілки кінетичного графа приписується чисельне значення - вага гілки, який відповідає константі швидкості реакції а ,, (табл. 13.1).
Вершини кінетичного графа відповідають станам системи 1-6 і з'єднані відповідно з можливими переходами (перший перехід 1 »2, другий перехід 2» 3 і т. Д.). Кожній лінії (дузі) на графі відповідає свою вагу. визначається кінетичними коефіцієнтами ау.
Таким чином, для повного кількісного опису динаміки переходів необхідно вирішити цю систему шести звичайних диференціальних рівнянь:
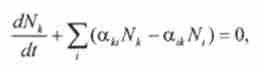
де Л / * (/), N, (t) - заселеності станів саркомерного містка.
У загальному випадку рішення такого роду можуть бути отримані з використанням чисельних методів.
Стаціонарні стану описують рівновагу між усіма сумами переходів як в одному, так і в іншому напрямку (замикання і розмикання сар- комостіка).
Розрахунок питомої потужності поперечно-смугастої м'язи. Відомо, що витрати енергії визначаються кількістю глюкози C6N | 206, що надходить до м'язів. Ця глюкоза «згоряє» в міоцитах, утворюючи Н20 і ССЬ:
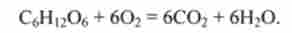
Витрата кисню може бути виміряний, а витрата глюкози можна розрахувати. Строго кажучи, в розрахунки повинен бути взнавств так званий дихальний коефіцієнт, проте в грубих оцінках цю деталь можна не враховувати. Коефіцієнт корисної дії становить приблизно 50%. Тоді неважко отримати, що 180 г глюкози відповідають енергії 2880 кДж. Звідси можуть бути визначені витрати на м'язову роботу.
Для розрахунку роботи саркомерного містка необхідно знати прикладену до містка силу і переміщення містка під дією цієї сили. Переміщення можна оцінити на основі молекулярної будови. Відома характерна довжина /ч згину «лапки» саркомера: це величина порядку /д ~ 10 нм (10 * м). Число елементарних актів стиснень «гачків» саркомера в результаті розпаду АТФ можна розрахувати, знаючи сумарну роботу AG '~ 30 кДж / моль і число Авогадро NA = 6-102я моль-1.
Звідси випливає, що робота за один цикл саркомерного містка приблизно дорівнює W = -AG7 NA «5-10 20Дж. Необхідно, крім того, зробити поправку на ефективність - врахувати ККД ~ 50%.
Отже, робота саркомерного містка становить W ~ 10 19 Дж. Тоді сила, прикладена до «лапці» саркомерного містка, оцінюється як F- Wlls = 10_n Н, що приблизно відповідає підйому маси 1 пг (1 (Г12 г).
Завдання розрахунку числа саркомерних містків м'язи формулюється так: визначити, яке число саркомерних містків потрібно для того, щоб за 1 з підняти на висоту 2 м вантаж масою 5 кг.
При виконанні таких порівняно невеликих навантажень може розвиватися потужність до 100 Вт. Один саркомерний місток протягом секунди робить приблизно 10 замикань з інтервалом в 1/10 с. Потужність для саркомерного містка становить величину порядку 10 19 Пн. Звідси неважко отримати, що число саркомерних містків, що здійснюють задану роботу, становитиме приблизно 1021.
З чим можна порівняти отримане число, багато це чи мало? Відомо, що в саркомере знаходиться близько кількох десятків «щіток» з рухомими зачепленнями - «гачками». На кожній такій «щітці» є приблизно 100 «гачків». Іншими словами, число цих «гачків» на один саркомер становить величину порядку 102. Кожен саркомерний місток має 10 «гачків». Тоді для здійснення зазначеної роботи буде потрібно приблизно 10,9саркомерів. Лінійні розміри саркомера такі: - 3 мкм у висоту і -10 мкм в діаметрі. Щільність м'язової тканини становить приблизно 1 г / мл. Після визначення обсягу (або маси) м'язи, здатної здійснити дану роботу, можна отримати значення питомої потужності: 1 Вт / кг. Це значення питомої потужності достатньо універсально. Воно характерно для енергетики м'яких тканин організму.
Таким чином, після нескладних оцінок, проведених на основі даних про біохімічний механізм переходів між станами, можна визначити «силу м'язи». Це значення сили задає масштаб біоадеквотного впливу технічних пристроїв БТС.
Якщо впливу (наприклад, ультразвукові або електромагнітні) на біосистему можуть привести до того, що до кожного саркомерному містку виявляється прикладеною сила близько 10 11 Н, то не виключена ймовірність ушкодження м'язового волокна. Цей простий, але важливий результат повинен враховуватися при розрахунку і конструюванні технічних пристроїв для тренувань - велоергометрія, різних тренажерів.
Питання до розд. 13.2