| Головна |
| «« | ЗМІСТ | »» |
|---|
Мейоз і запліднення забезпечують отримання організмами нового покоління еволюційно сформованого, збалансованого по дозам генів спадкового матеріалу, на основі якого здійснюється розвиток організму і окремих його клітин. Завдяки цим двом механізмам в ряду поколінь особин даного виду формуються певні видові характеристики і вид як реальна одиниця живої природи існує тривалий час. Однак у різних представників виду в силу постійно триваючого мутаційногопроцесу один і той же набір генів геному представлений різними їх алелями. Так як при статевому розмноженні у багатьох видів в вос- творі потомства беруть участь дві особини, то абсолютно очевидно, що в результаті запліднення різні зиготи отримують неоднаковий набір алелей в їх генотипах. Збільшенню генотипичного різноманітності представників виду сприяють також механізми, що приводять до переком- бинации батьківських алелей особини в її гаметах. Дійсно, якби гамети, що утворюються організмом, були однакові по набору алелей в їх геномі, то у нащадків однієї пари організмів при раздельнополости або одного гермафродитного організму не спостерігалося б генотипичного різноманітності. У кожному новому поколінні виду генотипично різними були б лише діти різних батьків.
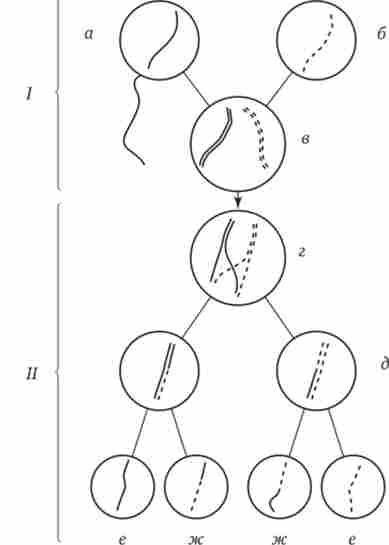
Мал. 3.72. Кроссинговер як джерело генетичного різноманіття гамет.
/ - запліднення батьківських гамет а й б з утворенням зиготи в; II - гаметогенезу в організмі, розвиненому з зиготи в; г - кросинговер, який відбувається між гомологами в профазі I; б - клітини, що утворилися після 1-го мейотичного поділу; е, ж - клітини, що утворилися після 2-го поділу мейозу (е - некроссоверние гамети з вихідними батьківськими хромосомами; ж - кроссоверние гамети з пере- комбінацією спадкового матеріалу в гомологічних хромосомах)
Реально в природі спостерігається різноманітність нащадків одних і тих же батьків. Наприклад, рідні брати і сестри розрізняються не тільки по підлозі, але і за іншими ознаками. Такі відмінності нащадків пояснюються тим, що в кожному акті запліднення зустрічаються генетично розрізняються гамети. Механізмом, що забезпечує різноманітність гамет, утворених одним і тим самим організмом, є мейоз, в ході якого відбувається не тільки зменшення вдвічі спадкового матеріалу, що потрапляє в гамети, а й ефективний перерозподіл батьківських алелей між гаметамі. Процесами, що приводять до перекомбінації генів і цілих хромосом в статевих клітинах, є кроссинговер і розбіжність бівалентов в анафазе I мейозу (див. Гл. 5).
Кроссинговер відбувається в профазі I мейозу в той час, коли гомологічні хромосоми тісно зближені в результаті кон'югації і утворюють біваленти. В ході кросинговеру здійснюється обмін відповідними ділянками між взаємно переплітаються хроматидами гомологічниххромосом (рис. 3.72). Цей процес забезпечує перекомбінація батьківських і материнських алелів генів в кожній групі зчеплення. У різних попередниках гамет кроссинговер відбувається в різних ділянках хромосом, в результаті чого утворюється велика різноманітність поєднань батьківських алелей в хромосомах.
Зрозуміло, що кросинговер як механізм рекомбінації ефективний лише в тому випадку, коли відповідні гени батька або матері хромосом представлені різними алелями. Абсолютно ідентичні групи зчеплення при кроссинговере не дають нових поєднань алелей.
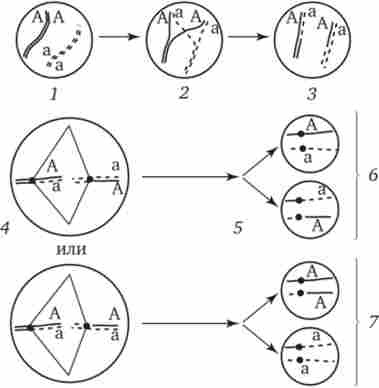
Мал. 3.73. Кроссинговер в соматичних клітинах.
1 - соматична клітина, в гомологічних хромосомах якої ген А представлений двома різними алелями (А і а); 2 - кросинговер; 3 - результат обміну відповідними ділянками між гомологічними хромосомами; 4- розташування гомологів в площині екватора веретена поділу в метафазі мітозу (два варіанти); 5 - утворення дочірніх клітин; 6 - освіту гетерозиготних по гену А клітин, схожих з материнською клітиною по набору алелей (Аа); 7-освіту гомозиготних по гену А клітин, що відрізняються від материнської клітини по набору алелей (АА або аа)
Кроссинговер відбувається не тільки в попередницях статевих клітин при мейозі. Він спостерігається також в соматичних клітинах при мітозі. Соматичний кроссинговер описаний у дрозофіли, у деяких видів цвілі. Він здійснюється в ході мітозу між гомологічними хромосомами, проте його частота в 10 000 разів менше частоти мейоті- чеського кросинговеру, від механізму якого він нічим не відрізняється.
В результаті мітотичного кросинговеру з'являються клони соматичних клітин, що розрізняються за змістом в них алелей окремих генів. Якщо в генотипі зиготи даний ген представлений двома різними алелями, то в результаті соматичного кросинговеру можуть з'явитися клітини з однаковими або батьківськими, або материнськими алелями даного гена (рис. 3.73).
У метафазі I мейозу в екваторіальній площині ахроматівного веретена шикуються біваленти, що складаються з однієї боку батька або однієї материнської хромосоми. Розбіжність гомологів, які несуть різний набір алелей генів в анафазе I мейозу, призводить до утворення гамет, що відрізняються по аллельному складу окремих груп зчеплення (рис. 3.74).
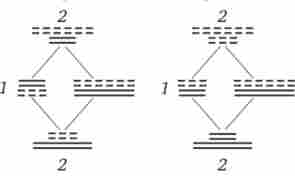
Мал. 3.75. Випадковий характер розташування бівалентов в метафазі (1) І незалежне розбіжність їх в анафазе (2) першого мейотіче- ського поділу
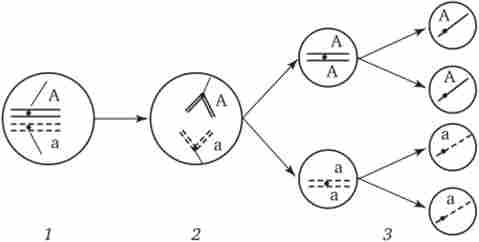
Мал. 3.74. Розбіжність гомологічниххромосом в анафазе I мейозу як джерело генетичного різноманіття гамет.
1 - метафаза I мейозу (розташування бивалента в площині екватора веретена поділу); 2 - анафаза I мейозу (розбіжність гомологів, які несуть різні алелі гена А до різних полюсів); 3 - другий мейотичний поділ (утворення двох типів гамет, що розрізняються по аллелям гена А)
У зв'язку з тим що орієнтація бівалентов по відношенню до полюсів веретена в метафазі I виявляється випадковою, в анафазе I мейозу в кожному окремому випадку до різних полюсів прямує гаплоїдний набір хромосом, що містить оригінальну комбінацію батьківських груп зчеплення (рис. 3.75). Різноманітність гамет, обумовлене незалежною поведінкою бівалентов, тим більше, чим більше груп зчеплення в геномі даного виду. Воно може бути виражено формулою 2 ", де п - число хромосом в гаплоидном наборі. Так, у дрозофіли п = 4, і кількість типів гамет, що забезпечується перекомбинацией батьківських хромосом в них, так само 24 = 16. У людини п = 23, і різноманітність гамет, обумовлене цим механізмом, відповідає 223, або 8 388 608.
Кроссинговер і процес розбіжності бівалентов в анафазе I мейозу забезпечують ефективну рекомбінацію алелей і груп зчеплення генів у гаметах, утворених одним організмом.
Запліднення. Випадкова зустріч різних гамет при заплідненні призводить до того, що серед особин виду практично неможливо поява двох генотипів однакових організмів. Що досягається за допомогою описаних процесів генотипичне різноманітність особин передбачає спадкові відмінності між ними на базі загального видового генома.
Таким чином, геном як вищий рівень організації спадкового матеріалу завдяки мейозу і запліднення зберігає свої видові характеристики. Але одночасно ці ж процеси забезпечують індивідуальні спадкові відмінності особин, в основі яких лежить рекомбінація генів і хромосом, т. Е. Ком бінатівную мінливість. Комбинативная мінливість, що виявляється в генотипичному різноманітності особин, підвищує виживаність виду в умовах, що змінюються його існування.