| Головна |
| «« | ЗМІСТ | »» |
|---|
Звільнені від транспортних плазматичних білків гормони щитовидної залози - три- йодтироніни і тироксин - відносно вільно проникають всередину клітин. У цитоплазмі тироксин, перетворюючись в трійодті- ронин, зв'язується зі специфічними ядерними рецепторами, які розташовуються на молекулі ДНК. Зв'язування гормон- рецепторного комплексу залежить як від іонного оточення, так і від особливостей первинної та вторинної структури акцепторной ДНК. Функціональне поєднання гормонорецепторного комплексу з ДНК залежить від гістонів, здатних підвищити спорідненість рецепторів до гормону і спорідненість комплексу до специфічних локусам ДНК, що і забезпечує виборчу регуляцію синтезу відповідних функціональних мРНК і рРНК. Під час трансляції індуковані РНК змінюють рівень синтезу кодованих ними білків.
На відміну від стероїдних і тиреоїдних гормонів білково-пептидні гормони і катехоламіни діють на гормонозалежні клітини через поверхневі рецептори, локалізовані в плазматичних мембранах. Виявлено три самостійних шляху проходження інформаційного гормонального сигналу з поверхні клітини: аденілатціклазную; кальцієвий; протеазний. Ці шляхи можуть реалізуватися самостійно, однак в реальних фізіологічних умовах вони тісно пов'язані.
Аденілатціклазную механізм реалізується за допомогою взаємодії гормонорецепторного комплексу з локалізованим в мембрані ферментом аденилатциклазой, утворюючим внутрішньоклітинний посередник цАМФ. Діяльність аденілатциклази здійснюється за участю чинників трансмембранного сполучення: гуанозинтрифосфат (ГТФ) - зв'язує N-білок, локалізований в ліпідному шарі мембрани; ГТФ, катіони магнію і марганцю і аніони заліза, компоненти цитоскелету (мікротрубочки і мікрофіламенти); кислі ліпіди мембран і деякі ферменти складають аденілатціклаз- ву систему. ГТФ, який грає основну роль в трансмембранному сполученні, пов'язує N-білок, і цей комплекс активує аде- нілатціклазу, яка здатна при утворенні одного гормон- рецепторного комплексу утворити до 500 молекул цАМФ. У комплексі з рецептором і аденилатциклазой N-білок здатний приєднувати до себе ГТФазу, яка, розщеплюючи ГТФ, інактівіруя N-білок і аденилатциклазу, припиняє процес активації цик- Лазні ферменту. Внутрішньоклітинна концентрація посередника 3 ', 5'-мононуклеотида (цАМФ) регулюється цитоплазматичним ферментом фосфодіестеразою, розщеплює його до неактивного 5'-мононуклеотида (рис. 12.4).
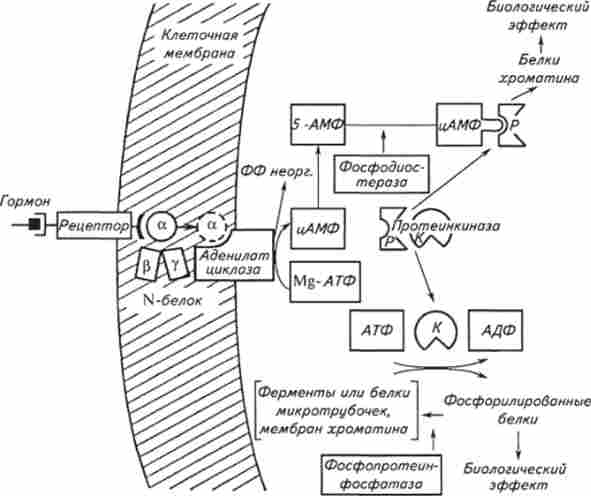
Мал. 12.4. Схема механізму дії гормонів у тварин за участю цАМФ
Утворився в клітці цАМФ специфічно взаємодієте регуляторними субодиницями цАМФ-залежних протеїн кіназ, відбувається дисоціація молекул ферменту на мономери. В результаті каталітична субодиниця активізується і забезпечує в цитоплазмі фосфорилирование цілого ряду синтезованих функціональних і структурних білків, змінюючи їх функціональний стан: активуються киназа фосфорілази, ліпаза, фосфоліпаза, Фосфопротеіни рибосом, гістони та т. Д. Крім того, комплекс цАМФ з однією з регуляторних субодиниць транслоціруется в клітинне ядро і надає виборче вплив на активність геному в ході транскрипції.
Необхідно відзначити, що стимулюючі ефекти гормонів можуть вдруге посилюватися і поширюватися за рахунок утворення простагландинів - похідних арахідонової кислоти, яка під впливом циклооксигенази перетворюється в циклічні ендоперекісі PgG2 і PgH2, з яких потім утворюються про- простагландинів. Простагландини, виходячи з материнської клітини, можуть діяти на ті ж або сусідні клітини і також стимулювати аденилатциклазную систему.