| Головна |
| «« | ЗМІСТ | »» |
|---|
Щитовидна залоза - одна з найбільших ендокринних залоз, розташовується в нижній частині гортані і верхньої частини трахеї у вигляді двох лопатей, з'єднаних між собою перешийком, що лежить на вентральній поверхні трахеї. Зовні покрита сполучнотканинною капсулою, в товщі якої розташовуються дрібні судини, капіляри, а також міелінізірованние і неміе- лінізірованние нервові волокна. Від капсули всередину залози відходять сполучнотканинні тяжі (септи), за якими проходять кровоносні і лімфатичні судини, нервові волокна. Паренхіма органу включає в себе кілька типів клітин: тиреоїдні
(Що формують стінку фолікула), парафоллікулярние і інтерфоллікулярние.
Фолікулярний епітелій складається в основному з паренхімних клітин і утворює основну структурно-функціональну одиницю щитовидної залози - фолікул (Округлої або овальної форми пляшечку). Між фолікулами знаходяться прошарки сполучної тканини, пронизані кровоносними капілярами. Просвіт фолікула заповнений колоїдів, який безпосередньо прилягає до апікальної поверхні тиреоцитах з численними микроворсинками, освіченими виступами плазматичноїмембрани. Протилежна сторона тиреоцитах тісно контактує з перікапіллярним простором, причому в базальному відділі клітини плазматична мембрана утворює глибокі складки, що значно збільшує її поверхню, що контактує з кровоносною системою. Бічні поверхні тиреоцитов з'єднуються за рахунок замикальних комплексів, що надійно ізолює внутрішній простір фолікула від навколишньої сполучної тканини. У цитоплазмі тиреоцитов особливо сильно розвинений шорсткий ендоплазматичнийретикулум з численних канальців і більших порожнин, заповнених гомогенним дрібнозернистим матеріалом. Комплекс Гольджі дуже добре виражений і представлений трьома основними компонентами: великими вакуолями, сплощеними цистернами (ламелами) і мікробульбашками. У зоні комплексу Гольджі виявляються секреторні гранули різної величини, електронної щільності і форми, оточені мембраною. Крім характерних гранул в апікальній частині цитоплазми виявляються колоїдні краплі, оточені мембраною.
Вихідними продуктами для біосинтезу йодованих гормонів служать амінокислота тіроніна і йодид. Що поставляється з кров'ю йодид в тиреоцитах, піддаючись окислення, перетворюється в молекулярний йод, який вступає в з'єднання з тіроніна, і через ряд проміжних сполук (монойодтіронін, дійод- тіроніна) утворюються тироксин (Тетрайодтіронін) і трійодтіро- нин. Вважається, що ці гормони синтезуються в вільному стані в цитоплазмі фолікулярнихклітин і необхідне для організму кількість гормону відразу після освіти надходить в кров, а його надлишок включається в молекули тиреоглобуліну і накопичується в колоїді. тиреоглобулін - велика глікопротеі- нова молекула, синтезована по загальній схемі синтезу білка. Звільнення тиреоглобуліну з секреторних гранул в порожнину фолікула відбувається або шляхом дифузії через обмежує мембрану, або шляхом екзоцитозу. Іодірованіе тиреоглобуліну відбувається в основному в просвіті фолікула і на кордоні між колоїдів і апікальної мембраною. При зниженні потреби організму в тиреоїдних гормонах внутрішньоклітинний гідроліз тиреоглобуліну припиняється і білок виводиться в порожнину фолікула, де накопичується у вигляді гормонального резерву. Накопичений в порожнині фолікула тіреоглобу- лин надходить в цитоплазму тиреоцитах за рахунок мікропіноцитоз колоїду: при підвищеній потребі можливо загарбання великих порцій за допомогою псевдоподий. Утворилися в результаті ендоцитозу колоїдні краплі поступово переміщуються до базальної частини клітини; при зустрічі з лізосомами відбуваються їх тісний контакт і злиття, причому гідролітичні ферменти лізосом надходять всередину колоїдних крапель. Колоїдні краплі зменшуються, а надійшли в цитоплазму тиреоїднігормони дифундують в перікапіллярним простір і потім проникають в кровоносне русло. При виснаженні запасів колоїду в порожнині фолікула синтез, йодування і протеолітичну розщеплення тиреоглобуліну повністю здійснюються в цитоплазмі тиреоцитов (рис. 12.9).
Трийодтиронин неміцно пов'язаний з білками крові і швидко проникає в клітини, а у тироксину цей зв'язок міцніше, тому він здатний довго циркулювати в кровоносній руслі. Фізіологічним дією в основному володіє трийодтиронін, так як він в 5 ... 6 разів активніше тироксину і, проникаючи в клітини тканин-мішеней, впливає на ферментні системи, локалізовані в мітохондріях (ферменти трикарбонових циклу, фосфорилювання, залізовмісні дихальні та ін.). Гормони шітовідной залози регулюють процеси росту і розвитку, величину енергетичного обміну, білковий, вуглеводний, жировий, водний і мінеральний обмін, тим самим впливаючи на діяльність серця, нервову систему і статеві залози.
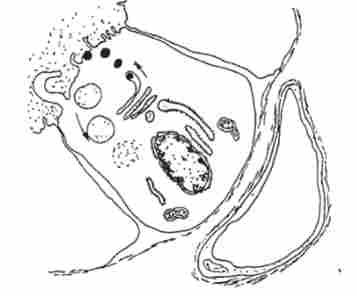
Мал. 12.9. Схема секреторного процесу в тіреоціте
Демонстративним прикладом участі тиреоїдних гормонів в регуляції росту і розвитку служить вплив щитовидної залози на метаморфоз амфібій. При видаленні щитовидної залози пуголовки ніколи не перетворяться на дорослу форму, виростаючи, залишаються на стадії личинки, а згодовування щитовидної залози або введення на її основі препаратів прискорюють метаморфоз амблістоми (личинкової форми аксолотлів) та інших земноводних. У птахів і ссавців гормони щитовидної залози відіграють важливу роль під час ембріонального розвитку. При виключенні функції залози у птахів гальмується ріст тіла, затримуються процеси окостеніння в кінцівках, порушується процес ембріонального оперення і т. Д. З активністю щитовидної залози пов'язані і видові особливості розвитку птахів. Так, у виводкових видів пташенята відразу після вилуплення з яйця здатні бігати і самостійно харчуватися, так як щитовидна залоза активно функціонує вже на ранніх стадіях зародкового розвитку. У птенцових видів, пташенята яких вилуплюються сліпими і безпорадними, щитовидна залоза в ембріональний період розвинена слабо і її активність зростає тільки після вилуплення з яйця.
Аналогічна закономірність виявляється і у ссавців - зрелорожденние дитинчата (здатні самостійно пересуватися) мають щитовидну залозу, активно функціонує на ранніх стадіях ембріонального розвитку. У ссавців, дитинчата яких з'являються на світ сліпими і голими, діяльність щитовидної залози проявляється незадовго до народження. Слід, однак, враховувати, що деяка кількість тиреоїдних гормонів потомство отримує з організму матері через плаценту і з молоком. Гормони щитовидної залози істотно впливають на розвиток кісткової тканини. При видаленні щитовидної залози за рахунок дії соматотропного гормону посилюється зростання трубчастих кісток, стимулюється проліферація епіфізарних хрящів, але затримується диференціація кісткової тканини. Введення тироксину гіпофізектомірованних тваринам викликає окостеніння епіфізів при незначному зростанні кісток. Зростання і прорізування зубів у ссавців залежать від вмісту в крові тиреоїдних гормонів. Тиреоїдні гормони стимулюють регенерацію тканин і сприяють загоєнню ран: грануляційна тканина швидше заповнює дно рани, і епітелізація йде більш інтенсивно.
Вплив щитовидної залози яскраво проявляється при оцінці основного обміну: тіреоідектомія знижує його на 45 ... 59%. Підвищення змісту гормонів за рахунок введення тироксину збільшує рівень основного обміну до 50% в порівнянні з нормою і вихід енергії при виконанні механічної роботи. Гормони щитовидної залози різко посилюють окисні процеси і теплопродукція, причому цей ефект виявляється і на ізольованих органах. Інкубіруемой фрагменти м'язів, печінки і нирок від тварин, яким попередньо вводили тироксин, споживають значно більше кисню, ніж аналогічні тканини, взяті у тіреоідектомірованних тварин. In vivo при гіпофункції щитовидної залози у тварин порушується терморегуляція, знижується температура тіла і вони погано переносять охолодження.
Гормони шітовідной залози здатні значно підвищити потребу організму в метаболітах: посилюється витрачання білка і жиру, баланс азоту стає різко негативним, азот у складі сечовини виводиться з сечею. При гіперфункції щитовидної залози зростає інтенсивність основного обміну: виявляється різке схуднення, витрачається до 70% запасів жирових депо, вміст холестерину в крові знижується. Вуглеводний обмін також інтенсифікується: рівень глюкози в крові дещо підвищується, а печінку швидко втрачає запаси глікогену. В основі фізіологічної дії тиреоїдних гормонів лежить регуляція інтенсивності дихання клітин, їх безпосередній вплив на поглинання кисню мітохондріями і іншими структурними компонентами клітин, посилення окислювальних реакцій і подальша зміна активності ряду ферментів і вплив на ядерний апарат клітин. Тироксин сприяє переносу амінокислот до рибосом, причому максимальне використання резорбироваться амінокислот відбувається в тому випадку, коли всі вони присутні в клітці відповідно до вимог певного кількісного співвідношення і послідовності інформаційної РНК.
Розвиток і функціональний стан нервової системи знаходяться під постійним впливом щитовидної залози. Її видалення призводить до порушення розвитку мозку: нейрони мають менші розміри, затримується миелинизация нервових волокон. Повноцінна диференціювання мозку, що розвивається відбувається при достатньому рівні тиреоїдних гормонів в організмі. В ході диференціювання і дозрівання нервової клітини існує критичний період, під час якого гормони роблять свій вплив на морфогенез, тому порушення функцій щитовидної залози в ранньому віці призводить до кретинізму. Надалі вони необхідні для підтримки метаболізму і функціональної активності нейрона. Тіреоідекто- мія різко знижує збудливість тварин і послаблює у них активні і пасивні оборонні реакції, пригнічує діяльність вищих нервових центрів. При гіпотиреозі умовні рефлекси виробляються з великими труднощами і вимагають постійного підкріплення: введення тиреоидина відновлює рефлекторну діяльність мозку.
Гормони щитовидної залози істотно впливають на діяльність серця, змінюючи ритм його скорочень. При видаленні щитовидної залози скорочення серця урежаются, а при введенні тироксину інтактним тваринам частота серцевих скорочень підвищується в 1,5 рази. Передозування гормону може привести до тріпотіння передсердь. При функціональних гіпотиреоїдних станах, характерних для зимнеспящих тварин (ведмедів, їжаків, бабаків, кажанів), спостерігається зниження температури тіла, зменшення рівня основного обміну і суттєве уповільнення ритму серцевих скорочень. Періоду пробудження у цих тварин передує активізація діяльності щитовидної залози; введенням тироксину можна припинити зимову сплячку і забезпечити пробудження у їжаків. Фізіологічний ги пертіреоз відзначається при функціональних станах тварин, які вимагають інтенсифікації обміну речовин: вагітність, лактація (особливо у високопродуктивних тварин).
Тиреоїдні гормони прискорюють всмоктування глюкози в шлунково-кишковому тракті, беруть участь в регуляції рівня глікогену в печінці, посилюють секрецію молока і збільшують вміст у ньому жиру. Під дією тиреоїдних гормонів різко прискорюються процеси окислення жирних кислот в мітохондріях.
Регуляція секреції тиреоїдних гормонів здійснюється перш за все за рахунок виділення специфічного стимулюючого речовини - тиреотропного гормону передньої долі гіпофіза. У гіпофізектомірованних тварин зменшуються розмір і маса щитовидної залози, розвивається атрофія тканини і в першу чергу редукуються новостворені фолікули на тлі симптомів тиреоїдної недостатності. Підвищення секреції тиреотропного гормону збільшує продукцію і надходження в кров гормонів щитовидної залози. Але збільшення концентрації тиреоїдних гормонів позначається на продукції тиролиберина в гіпоталамусі: надлишок гормону гальмує тиреотропну функцію гіпофіза, що веде до зниження секреції тироксину в щитовидній залозі і реалізації зворотного негативного зв'язку, що забезпечує в нормі сталість гормонального рівня.
Поряд з системою гуморальної регуляції, в якій тірео- тропний функцій гіпофіза корелюється з виділенням АКТГ і соматотропного гормону, активність щитовидної залози істотно залежить від функціонального стану і енергетичних потреб організму. При охолодженні, що викликає роздратування холодових рецепторів, рефлекторно посилюється секреція тиреотропного гормонів, обумовлена впливом гіпоталамічного тиролиберина. Разом з впливом гіпоталамуса регулюючий вплив на стан щитовидної залози надає вегетативна нервова система: симпатичний відділ посилює активність органу, а парасимпатичний пригнічує. Впливом симпатичного відділу вегетативної нервової системи пояснюється прискорене виділення тиреоїдних гормонів в умовах стресових ситуацій, що забезпечує пристосування організму до несприятливих факторів середовища: охолодження, негативним емоціям і т. Д.
Найбільш часто порушення діяльності щитовидної залози викликане недостатнім надходженням йоду в організм. Зниження рівня тиреоїдних гормонів позначається на діяльності гіпоталамуса і гіпофіза, який відповідає посиленою секрецією тиреотропина, що викликає розростання тканини щитовидної залози (зоб). Збільшення залози може бути за рахунок розростання сполучної тканини (простий зоб) або збільшення обсягу фолікулярної тканини (колоїдний зоб). У місцевостях, де грунт і вода бідні йодом (гірські райони, зони підзолистих, сірих лісових і солонцевих ґрунтів), можливий розвиток зобної ендемії.
Недостатнє надходження йоду в організм призводить до зниження освіти тиреоглобуліну, що володіє властивостями каротінази (ферменту, відповідального за перетворення каротину в вітамін А). Таким чином, надлишок тиреоїдних гормонів в тканинах організму нейтралізується вітаміном А і у випадках гіпертиреозу можливий А-авітаміноз. Надходження вітаміну А або каротину з кормом нормалізує основний обмін і піднімає рівень глікогену в печінці. На активність щитовидної залози впливають повноцінність білків, жирів, вуглеводів і наявність антитиреоїдних речовин гойтро- генів (Містяться в соєвих бобах, горосі, білому конюшині, капусті і турнепсі), які вибірково порушують процеси зв'язування йоду в щитовидній залозі. Недолік тирозину і фенілаланіну, надлишок рослинних жирів і тривалий, одноманітне вуглеводне харчування призводять до розвитку дистрофічних змін в залозі і порушення продукції тиреоїдних гормонів.
Парафоллікулярние клітини (К-клітини) розташовуються в інтерфоллікулярних острівцях і стінці фолікулів щитовидної залози. Вони суттєво відрізняються за своєю структурою від тиреоцитов: мають низьку електронну щільність (світлі клітини) і містять в цитоплазмі велику кількість секреторних гранул і везикул. К-клітини не контактують з колоїдом і максимально наближені до капілярів в інтерфоллікулярних острівцях; розташовуються на периферії і на зовнішній поверхні фолікулів. Гранули, локалізовані в цитоплазмі К-клітин, мають діаметр 200 ... 280 нм, містять тонко гранульований матеріал і оточені одиночної мембраною. Поблизу плазматичноїмембрани виявляються ознаки екзоцитозу - злиття мембрани секреторною гранули з плазма- леммой і звільнення її вмісту в пери капілярний простір. Встановлено, що К-клітини продукують тіреокаль- цітонін - поліпептид, що складається з 32 амінокислот, що викликає падіння рівня кальцію і фосфору в крові. Це зниження концентрації кальцію в циркулюючої крові обумовлено гальмуванням його мобілізації з кісток, причому істотно змінюється діяльність клітин кісткової тканини: пригнічуються остеокласти і активуються остеобласти, що призводить до фіксації кальцію і фосфору. Поряд з цим суттєво збільшується видалення фосфору з сечею. Тиреокальцитонін посилено секреті- руется і надходить у кров при підвищенні концентрації кальцію, що є важливою ланкою в підтримці сталості його рівня у внутрішньому середовищі організму.
Околощітовідние (паращитовидні) залози. У ссавців представляють собою парні освіти і розташовані близько щитовидної або вилочкової залози або занурені в їх тканину. Ці невеликі округлі залози оточені власної сполучнотканинної капсулою. Від капсули в товщу органу відходять тонкі септи, між якими знаходяться тяжі і групи залізистих клітин - паратіреоцітов. Паратіреоціти розташовуються радіально навколо численних капілярів, які пронизують тіло органу. Структурна організація клітин паренхіми близько-щитовидної залози носить характерні ознаки інтенсивного секреторного процесу: розвиненою ендоплазматичнийретикулум представлений численними сплощеними канальцами і сво- боднолежащімі рибосомами, які зумовлюють дрібнозернисту структуру цитоплазми; комплекс Гольджі добре розвинений і представлений великою кількістю мікропухирців, що мають дрібнозернистий вміст.
Крім мікропухирців в цитоплазмі паратіреоцітов виявляються більші (діаметр 150 ... 200 нм) гранули, що формуються в комплексі Гольджі і виділяються з клітин в області латеральних клітинних контактів. Поряд з секреторними структурами паратіреоціти можуть містити ліпідні краплі, ліпопігментние тільця і лізосомальні освіти. Крім цих клітин, що описуються при світловій мікроскопії як «головні», паренхіма околошітовідних залоз містить електроноплотние, або «оксифільні», клітини, функціональне значення яких ще не встановлено. Не виключена можливість того, що околощітовідние залози утворені клітинами одного типу (головними клітинами), цитоплазма яких може змінюватися в широких межах залежно від функціонального стану. Паратіреоціти контактують з одним або декількома капілярами, причому періендотеліальное простір розширений і містить крім двох базальних мембран і спеціальні клітини судинного оточення - перицитам. Ендотелій капілярів уплощен і має численні фенестри.
Якщо у тварини видалити околощітовідние залози (параті- реоідектомія), то на 2 ... 3-е добу порушується збудливість нервової системи: хода стає більш напруженою, з'являється фібрилярні посмикування окремих м'язів голови і тулуба. Поодинокі скорочення посилюються і переходять в бурхливі напади загальних судом (титания), які повторюються все частіше. Тварина незабаром гине за рахунок зниження вмісту кальцію в крові, оскільки певний рівень кальцію необхідний для перебігу найважливіших життєвих процесів. Внутрішньовенне введення хлористого кальцію оперованих тварин здатне тимчасово припинити судоми і знизити збудливість ЦНС.
Околошітовідние залози секретують гормон - паратгор- мон, поліпептид з молекулярною масою близько 8500, що складається з 83 амінокислотних залишків. Паратгормон регулює концентрацію кальцію і фосфору. В результаті його дії підвищується рівень кальцію в крові і тканинах організму, а вміст фосфатів знижується. Виходячи з того, що кальцій потрапляє в організм з їжею і надходить у кров у тонкому кишечнику, дія гормону проявляється вже на рівні кальційтранспортірующіх систем - ентероцитів. Основним депо кальцію в організмі служить кісткова тканина, яка містить до 99 % Найбільше кальцію в складі гідрооксіапатіта.
Дія паратгормону проявляється в мобілізації запасеного в кістки кальцію. Посилюється діяльність руйнують кістку остеокластів і пригнічується функція фіксують кальцій остеобластів, відбувається деполимеризация мукополісаха- рідов основної речовини кістки, її декальцинація і надходження іонів кальцію і фосфорної кислоти в кров. Одночасно з цим зменшується реабсорбція фосфатів з сечі: вони швидко виводяться, і їх рівень в крові швидко знижується. Продукція паратгормону регулюється в основному рівнем кальцію в крові за принципом зворотного негативного зв'язку. У випадках високої потреби організму в кальції, що настає при вагітності і лактації, коли для плоду, що розвивається або освіти казеїну необхідне надходження додаткових значних кількостей кальцію, виявляється фізіологічна гіперфункція околощітовідной залози. Якщо не збільшити вміст кальцію в кормі, то можлива декальцинація (остеомаляція і остеопороз) кісткової тканини материнського організму.