| Головна |
| «« | ЗМІСТ | »» |
|---|
Історію розвитку поглядів на одиниці спадковості (гени), відкриті Г. Менделем, можна умовно розділити на кілька періодів. Відповідно до «класичної» точкою зору, яка побутувала в 30-і рр. XX ст., Ген розглядали як неподільну одиницю генетичної передачі, функції, мутації і рекомбінації. Починаючи з 1940-х рр. в зв'язку з встановленням генетичної ролі ДНК формується «неокласична» концепція, згідно з якою ген (цистрон) являє собою ділянку молекули ДНК із специфічною послідовністю нуклеотидів, що визначає первинну структуру синтезується молекули мРНК і відповідного поліпептиду або одиночної молекули тРНК або рРНК. При цьому ген підрозділяється на складові частини у вигляді елементарних одиниць мутації (Мутона) і рекомбінації (рекон), Які можуть бути ідентифіковані як певні ділянки полинуклеотида. Гени, що детермінують структуру поліпептидів і молекул РНК, отримали назву структурних генів. Сучасний період розуміння гена, що почався з 1970-х рр., Пов'язаний з появою нових знань про переривчастої ( «мозаїчної») структуру генів еукаріот і ряді інших особливостей генетичної організації різних організмів (перекриваються гени, що повторюються гени, псевдогени, мобільні гени і ін. ).
В рамках класичної (формальної) генетики прийнято розглядати ген як структурну одиницю, детерминирующую елементарна ознака (фен) організму. Сукупність усіх генів окремого організму (індивідуума) називають його генотипом, а сукупність успадкованих ознак - фенотипом. Терміном «геном» прийнято позначати сукупність всіх генетичних елементів (ДНК хромосом, мітохондрій, плазмід і ін.), Які є постійними для організмів даного виду. Слід зауважити, що розміри геномів (кількості геномної ДНК або РНК у відповідних вірусів) мають істотні відмінності у організмів, що відносяться до різних рівнів організації живої матерії (вірусів, бактерій, еукаріот).
Досягнення сучасного етапу у вивченні структури і функцій генів пов'язані з розробкою і широким використанням технологій генетичної інженерії, в тому числі методів клонування фрагментів ДНК (генів) різних організмів. Для молекулярного клонування можуть бути використані ферменти, що отримали назву рестрикційних ендонуклеаз (рестриктаз), які здатні розщеплювати ( «розрізати») специфічні нуклеотидні послідовності молекули ДНК з руйнуванням фосфодіефірних зв'язків і утворенням лінійних фрагментів молекули. В якості носіїв (векторів) клонованих генів зазвичай використовують невеликі кільцеві молекули ДНК вірусів або бактеріальних плазмід.
Прикладом може служити дію рестріктази EcoRl, яка здатна «дізнаватися» ділянки молекули ДНК, що містять шести нуклеотидні інвертовані послідовності (б'-ГААТТЦ-З 'на одній нитки і 3'-ЦТТААГ-5' на інший (комплементарної) нитки), і вносити розриви між нуклеотидами Г і А кожної з ниток молекули (рис. 5.24). Подальший поділ цих ниток призводить до появи однониткових ( «липких») решт утворилися фрагментів молекули, які, однак, можуть легко возз'єднуватися по комплементарному принципом за допомогою ферменту лігази, здатного відновити цілісну структуру молекули.
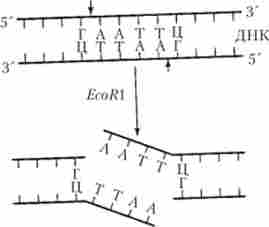
Мал. 5.24. Механізм дії рестріктази EcoR
На рис. 5.25 приведена схема експерименту з клонування одного з рестрикційних фрагментів хромосомної молекули ДНК, що містить потрібний досліднику структурний ген, в кільцеву векторну молекулу ДНК (плазміди), яка попередньо розрізається тієї ж рестриктазой, отже, має «липкі» кінці, необхідні для подальшого возз'єднання з клонують хромосомним фрагментом. Як вектора зазвичай підбирається така молекула ДНК, яка має лише один полінукпеотідний ділянку, впізнаваний використовуваної рестриктазой, т. Е. Один ділянку розрізання, тому в результаті її рестрикції утворюється один лінійний фрагмент, що має два комплементарних один одному «липких» кінця. Отриману таким способом гібридну молекулу ДНК (плазмідний вектор з включеним в його структуру хромосомних геном) можна потім ввести в бактеріальну клітину за допомогою трансформації і копіювати в процесі розмноження бактерій, які є господарями цієї молекули.
Подальше виділення копіювати ДНК клонованого гена, вирізаної тієї ж рестриктазой зі структури векторної молекули, дає можливість проводити детальний молекулярно-генетичний аналіз цього гена, включаючи визначення його нуклеотидної послідовності (секве- нирование гена). До теперішнього часу створені великі «бібліотеки» клонованих генів (клонотекі) різних організмів, які використовуються як для дослідницьких робіт, так і при вирішенні низки практичних завдань.
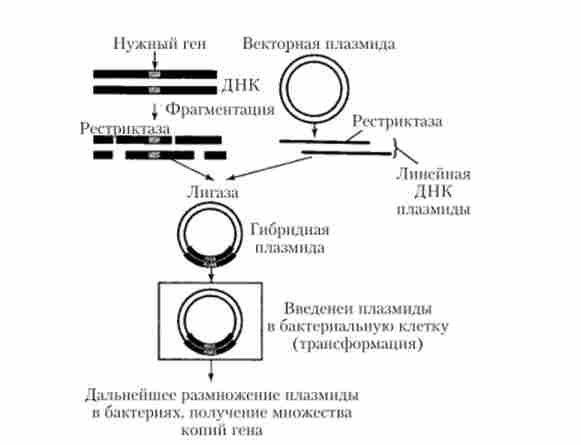
Мал. 5.25. Клонування гена, що знаходиться в структурі фрагмента хромосомної ДНК, в векторну молекулу плазмідної ДНК з використанням рестриктаз і лігази
Більшість структурних генів прокаріотів (бактерій) представлено безперервними ділянками молекули ДНК, вся інформація яких використовується при синтезі кодованих поліпептидних ланцюжків. Отже, генетична інформація прокариотичного гена реалізується повністю. У деяких дрібних вірусів була виявлена незвичайна структурно-функціональна організація генетичного матеріалу у формі перекриваються генів (за принципом «ген в гені»), яка дозволяє здійснювати ще більш економне використання наявних дуже обмежених інформаційних можливостей генома. Так, наприклад, деякі ділянки ДНК одного з найбільш дрібних бактеріофагів СРХ 174 (табл. 5.5) містять інформацію не одного, а одночасно двох різних генів, що дозволяє геному 'настільки малих розмірів кодувати не менше 9 різних білкових молекул. Зчитування інформації перекриваються генів починається з різних стартових точок однієї і тієї ж нуклеотидноїпослідовності, т. Е. Є різні рамки зчитування цієї послідовності.
Розміри геномів різних організмів
|
організми |
Число пар нуклеотидів гаплоидного генома |
|
бактеріофаг <рХ 174 |
5,4 х 103 |
|
бактеріофаг X |
5,0 х Ю4 |
|
бактеріофаг Т4 |
1,8 х Ю5 |
|
Кишкова паличка |
4,2 х Ю6 |
|
дріжджі |
1,8 х Ю7 |
|
нематода |
8,0 х Ю7 |
|
Людина |
3,0 х ю9 |
На відміну від прокаріотів для еукаріот типовим є переривчастий характер структурно-функціональної організації генів. Інформація такого гена про структуру синтезованого поліпептиду існує не у вигляді безперервної нуклеотидної послідовності певної ділянки молекули ДНК, а в формі кодують фрагментів (екзонів), які перериваються (поділяються) «інертними» нуклеотидними послідовностями (нітрон), котрі приймають прямої участі в кодуванні цього поліпептиду . Отже, гени різних еукаріотичних організмів є мозаїку з декількох чергуються в певному порядку екзонів і інтронів. Розміри интронов в складі таких генів коливаються від 10 до більш ніж 1000 пар нуклеотидів. Прикладом може служити мозаїчна структура генів, що кодують синтез поліпептід- них ланцюжків а- і р-глобіну (рис. 5.26), які формують тетрамерную структуру молекули гемоглобіну людини, що містить дві а- і дві р-ланцюжка. Зафарбовані ділянки (рис. 5.26) - райони генів, що кодують структуру поліпептидів (екзонів), які розділені интронами (незафарбовані ділянки). Цифри над генами вказують амінокислотні залишки кодованого поліпептиду (після сплайсингу). Заштриховані ділянки, які транскрибуються в мРНК, але не транслюються в білок, прийнято розглядати як нетрансльовані частини першого і останнього екзонів.
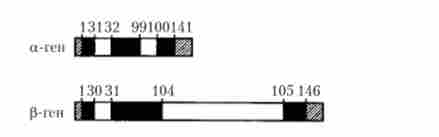
Мал. 5.26. Структура генів а- і р-глобіну людини
Передбачається, що нітроном можуть брати участь в регуляції процесингу РНК. Є дані, що дозволяють вважати, що вони, ймовірно, істотно впливають на процеси рекомбінації між гомологічними генами. Існує також гіпотеза про те, що по Інтрони ділянках відносно легко і часто можуть рекомбінуватися гени різних білків або гени, що детермінують білки одного сімейства, але накопичили різні мутації. Можна вважати, що такі властивості интронов повинні прискорювати еволюцію білкових молекул, полегшуючи процеси еволюції еукаріот в цілому, тим самим забезпечуючи їм значні переваги в порівнянні з прокариотами. Як «еволюційного резерву» еукаріот можна, ймовірно, розглядати і виявляються в їх геномах псевдогени, які представляють собою нуклеотидні послідовності ДНК, гомологічні послідовностям відомих (функціонуючих) генів, але з тих чи інших причин не виявляють інформаційної активності, т. Е. дають кінцевого зрілого продукту.
Однією з особливостей генетичної організації еукаріотів є також присутність в їх геномах значного числа повторюваних генів, що кодують первинну структуру тРНК, рРНК, білків-гістонів, а також інших (менш протяжних і не завжди ідентифікованих в плані функціональної значущості) повторюваних послідовностей ДНК, кількість копій яких може варіювати від одиниць до декількох тисяч і більше. Так, наприклад, в гаплоидном геномі людини, що містить близько 3 х 109 пар нуклеотидів, що повторюються послідовності ДНК складають приблизно 30%, тоді як решта 70% генома представлені «унікальними» послідовностями, які існують в одиничних копіях.
У геномах різних організмів (прокаріот і еукаріот) виявлені також мобільні (транспозіруемие) гени.