| Головна |
| «« | ЗМІСТ | »» |
|---|
Темнові реакції, пов'язані з асиміляцією С02 у зелених рослин, були детально вивчені за допомогою радіоізотопного аналізу роботами М. Кальвіна, А. А. Бенсона і Дж. А. Бассама і привели до створення загальної гіпотетичної схеми, відомої під назвою циклу Кальвіна-Бассама- Бенсона.
Центральної реакцією, що забезпечує перехід С02 в органічну форму, є реакція карбоксилирования і одночасного розщеплення рибулозо-1,5-дифосфата з утворенням двох молекул 3-фосфогліцерата (ЗФГ), яка каталізується ферментом рибулозо-1,5-діфосфаткарбоксілазой (РК):
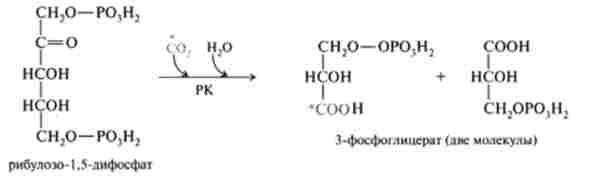
Фермент РК зустрічається тільки у фотосинтезуючих організмів і відсутній в тварин тканинах. Цей фермент має молекулярну масу 55 kDa і складається з 16 субодиниць двох типів: 8 - каталітичних і 8 - регулятор-
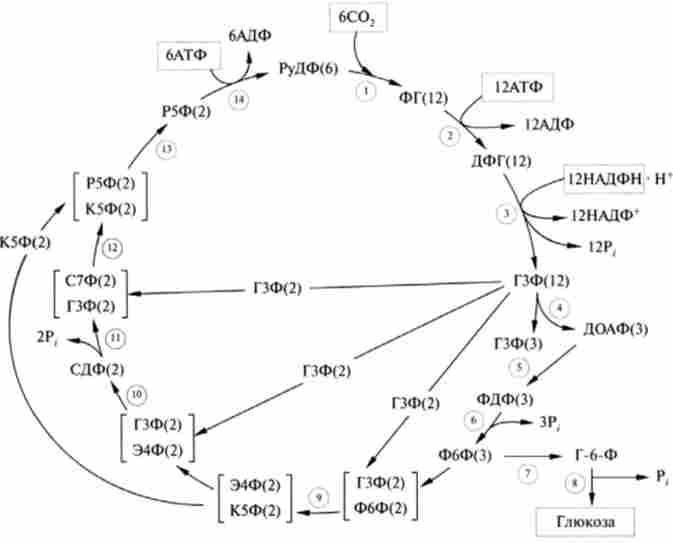
Мал. 16.4. Схема реакцій циклу Кальвіна:
цифри в гуртку позначають номери реакцій; в дужках наведено кількість молекул, що ілюструють стехіометрію перетворень; РуДФ - рибулозо-1,5-дифосфат; ФГ - 3-фосфогліцерат; ДФГ - 1,3-дифосфоглицерата; ГЗФ - глицеральдегид-3-фосфат; ДОАФ - дегідроксіаіетон-3- фосфат; ФДФ - фруктозо-1,6-дифосфат; Ф6Ф - фруктозо-6-фосфат; Г6Ф - глюкозо-6-фосфат; Е4Ф - зрітрозо-4-фосфат; К5Ф - ксилулозо-5-фосфат; С7Ф - седогеітулозо-7-фосфат; СДФ - седогептулозо-1,7-дифосфат; Р5Ф-рибоза-5-фосфат; Ру5Ф - рибулозо-5-фосфат
них. Для повного біосинтезу глюкози, т. Е. Освіти всіх її шести вуглецевих атомів з С02 в циклі Кальвіна, на кожну фіксовану молекулу діоксиду вуглецю необхідна одна молекула рібулозодіфосфат, яка регенерує в кінці циклу (рис. 16.4).
Реакції (2) - (8) циклу Кальвіна збігаються з такими в процесі глю- конеогенеза в тварин тканинах (гл. 20), за винятком того, що донором відновних еквівалентів в реакції (3) виступає НАДФН, а не НАДН. В ході цих реакцій 6 молекул С02 включаються в глюкозу. Реакції (9) - (14) спрямовані на регенерацію 6 молекул рибулозо-1,5-ліфосфата, який необхідний для того, щоб міг розпочатися новий оборот циклу Кальвіна. Цей процес досить складний і включає реакції, що каталізуються ферментом гліколізу альдолазой і ферментами пентозофосфатного шляху окислення глюкози транскетолази, епімерази і ізомерази. Хімізм реакцій наведено в гл. 18, схематично їх можна описати таким чином:
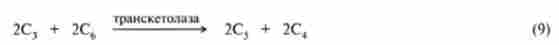
де С5 - ксилулозо-5-фосфат; З4 - еритроза-4-фосфат

де С7 - седогептулозо-1,7-дифосфат
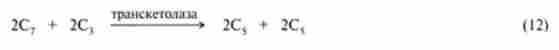
де одна пентоза - рибоза-5-фосфат; інша - ксилулозо-5-фосфат.
Продукти цих реакцій - шість молекул пентоз, з них чотири молекули ксилулозо-5-фосфату і дві молекули рибози-5-фосфату ізомеризуються при дії ферментів відповідно епімерази і ізомерази в шість молекул рибулозо-5-фосфату, які в заключній реакції циклу Кальвіна фосфорілізуются АТФ і перетворюються в шість молекул рибулозо-1,5-дифосфата. Сумарне рівняння фотосинтезу глюкози, з урахуванням регенерації ри- булозо-5-фосфату, можна записати таким рівнянням:
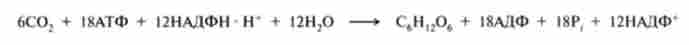
Таким чином, шість молекул С02 - це еквівалент однієї молекули глюкози, і, отже, на кожну асимілювати молекулу С02 витрачається три молекули АТФ і шість молекул НАДФН.
Регуляція темновой стадії фотосинтезу. Регуляторним ферментом перетворення С02 в вуглеводи є перший фермент циклу Кальвіна - ри- булозо-1,5-діфосфаткарбоксілаза. При регуляції по аллостерічному механізму інгібітором ферменту є один з центральних метаболітів циклу Кальвіна - фруктозо-1,6-дифосфат, а активатором - фруктозо-6-Фос фат. У свою чергу, обидва ефекту пов'язані з активацією циклу Кальвіна при дії квантів світла за схемою:
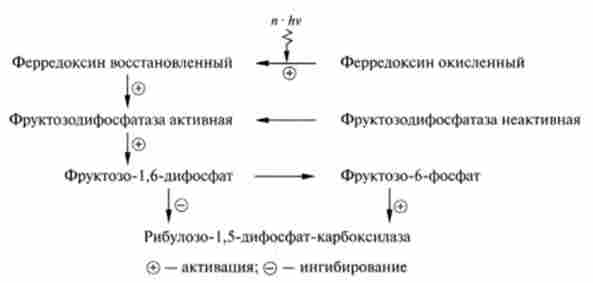
Таким чином, хоча карбоксилаза каталізує основну реакцію тем- нової фази, її активність опосередковано залежить від поглинання хлоропластами світла. Якщо освітлені хлоропласти синтезують АТФ і відновлюють НАДФ+, світло активує процес синтезу глюкози з С02, в ході якого використовуються продукти світлової фази.