| Головна |
| «« | ЗМІСТ | »» |
|---|
Довгий час ген розглядали як мінімальну частину спадкового матеріалу (геному), що забезпечує розвиток певної ознаки у організмів даного виду. Проте яким чином функціонує ген, залишалося неясним. У 1945 р Дж. Бідлом і Е. Татум була сформульована гіпотеза, яка вміщується у формулу «Один ген - один фермент». Відповідно до цієї гіпотези, кожна стадія метаболічного процесу, що призводить до утворення в організмі (клітці) якогось продукту, каталізується білком-ферментом, за синтез якого відповідає один ген.
Пізніше було показано, що багато білків мають четвертинних структуру, в утворенні якої беруть участь різні пептидні ланцюга. Наприклад, гемоглобін дорослої людини включає чотири глобінових ланцюга - 2а і 2 (3, які кодуються різними генами. Тому формула, що відображає зв'язок між геном і ознакою, була дещо перетворена: «Один ген - один поліпептид».
Вивчення хімічної організації спадкового матеріалу і процесу реалізації генетичної інформації призвело до формування уявлення про гені як про фрагмент молекули ДНК, транскрибується у вигляді молекули РНК, яка кодує амінокислотну послідовність пептиду або має самостійне значення (тРНК і рРНК).
Відкриття екзон-інтронів організації еукаріотичних генів і можливості альтернативного сплайсингу показали, що одна і та ж нуклеотидних послідовність первинного транскрипту може забезпечити синтез декількох поліпептидних ланцюгів з різними функціями або їх модифікованих аналогів. Наприклад, в мітохондріях дріжджів є ген box (або cob), що кодує дихальний фермент цитохром Ь. Він може існувати в двох формах (рис. 3.42). «Довгий» ген, що складається з 6400 п. Н., Має 6 екзонів загальною протяжністю 1155 п. Н. і 5 интронов. Коротка форма гена складається з 3300 п. Н. і має 2 інтрона. Вона фактично є позбавлений перших трьох інтронів «довгий» ген. Обидві форми гена однаково добре експресуються.
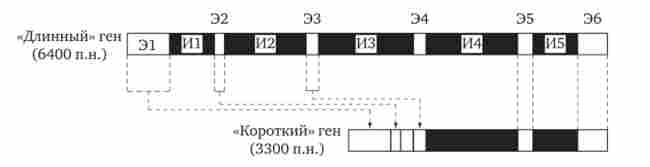
Мал. 3.42. Дві форми гена box (cob) мітохондрій дріжджів, що кодує цитохром Ь.
«Короткий» ген box утворюється в результаті видалення з «довгого» гена перших трьох інтронних ділянок і сплайсингу перших чотирьох екзонів; І - інтрон, Е - екзон; п. н.- пари нуклеотидів
Після видалення першого інтрони «довгого» гена box на основі об'єднаної нуклеотидноїпослідовності двох перших екзонів і частини нуклеотидів другого інтрона утворюється матриця для самостійного білка - РНК-матурази (рис. 3.43). Функцією РНК-матурази є забезпечення наступного етапу сплайсингу - видалення другого інтрона з первинного транскрипту і в кінцевому рахунку утворення матриці для цитохрому Ь.
Іншим прикладом може служити зміна схеми сплайсингу первинного транскрипту, що кодує структуру молекул антитіл у лімфоцитах. Мембранна форма антитіл має на С-кінці довгий «хвіст» амінокислот, який забезпечує фіксацію білка на мембрані. У секретируемой форми антитіл такого хвоста немає, що пояснюється видаленням в ході сплайсингу з первинного транскрипту кодують цю ділянку нуклеотидів.
У вірусів і бактерій описана ситуація, коли один ген може одночасно бути частиною іншого гена або деяка нуклеотидних послідовність ДНК може бути складовою частиною двох різних перекриваються генів. Наприклад, на фізичній карті генома фага ФХ174 (рис. 3.44) видно, що послідовність гена В розташовується всередині гена А, а ген Е є частиною послідовності гена D. Цією особливістю організації генома
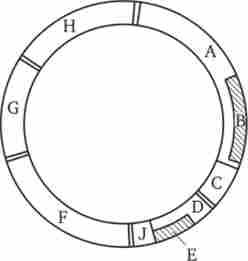
Мал. 3.44. Фізична карта геному фага ФХ174. Нуклеотидная послідовність гена В є одночасно частиною гена А, а ген Е становить частину гена D
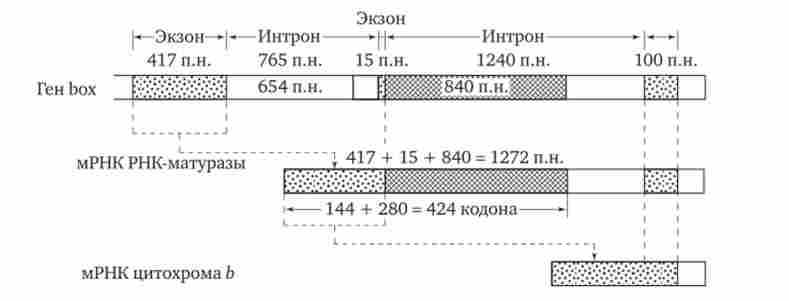
Мал. 3.43. Освіта матриці для РНК-матурази в ході сплайсингу первинного транскрипту гена цитохрому b мітохондрій дріжджів; п. н.- пари нуклеотидів
фага вдалося пояснити існуючу невідповідність між відносно невеликим його розміром (він складається з 5386 нуклеотидів) і числом амінокислотних залишків у всіх білках, що синтезуються, яке перевищує теоретично припустиме при даній ємності генома. Можливість складання різних пептидних ланцюгів на мРНК, синтезованої з перекриваються генів (А і В або Е і D), забезпечується наявністю всередині цієї мРНК ділянок зв'язування з рибосомами. Це дозволяє почати трансляцію іншого пептиду з нової точки відліку.
У геномі фага X були також виявлені перекриваються гени, що транслюються як із зсувом рамки, так і в тій же рамці зчитування. Передбачається також можливість транскрибування двох різних мРНК з обох комплементарних ланцюгів однієї ділянки ДНК. Це вимагає наявності промоторних областей, що визначають рух РНК-полімерази в різних напрямках уздовж молекули ДНК.
Описані ситуації, що свідчать про допустимість зчитування різної інформації з однієї і тієї ж послідовності ДНК, дозволяють припустити, що перекриваються гени являють собою досить поширений елемент організації генома вірусів і, можливо, прокаріотів. У еукаріот уривчастість генів також забезпечує можливість синтезу різноманітних пептидів на основі однієї і тієї ж послідовності ДНК.
Маючи на увазі все сказане, необхідно внести поправку в визначення гена. Очевидно, не можна більше говорити про гені як про безперервну послідовності ДНК, однозначно кодує певний білок. Мабуть, в даний час найбільш прийнятною все ж слід вважати формулу «Один ген - один поліпептид», хоча деякі автори пропонують її переінакшити: «Один поліпептид - один ген». У всякому разі, під терміном ген треба розуміти функціональну одиницю спадкового матеріалу, по хімічній природі що є полінуклеотидом і визначальну можливість синтезу поліпептидного ланцюга, тРНК або рРНК.