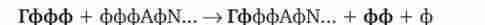
| Головна |
| «« | ЗМІСТ | »» |
|---|
За хімічною організації матеріалу спадковості і мінливості еукаріотичні і прокариотичні клітини принципово не відрізняються один від одного. Генетичний матеріал у них представлений ДНК. Загальним для них є і принцип запису генетичної інформації, а також генетичний код. Одні і ті ж амінокислоти шифруються у про- і еукаріот однаковими кодонами. Принципово однаковим чином у названих типів клітин здійснюється і використання спадкової інформації, що зберігається в ДНК. Спочатку вона транскрибується в нуклеотидну послідовність молекули мРНК, а потім транслюється в амінокислотну послідовність пептиду на рибосомах за участю тРНК. Однак деякі особливості організації спадкового матеріалу, що відрізняють еукаріотичні клітини від прокаріотичних, обумовлюють відмінності у використанні їх генетичної інформації.
Спадковий матеріал клітини прокаріотів міститься головним чином в єдиній кільцевої молекулі ДНК. Вона розташовується безпосередньо в цитоплазмі клітини, де також знаходяться необхідні для експресії генів тРНК і ферменти, частина з яких укладена в рибосомах. Гени прокаріот складаються цілком з кодують нуклеотидних послідовностей, що реалізуються в ході синтезу білків, тРНК або рРНК.
Спадковий матеріал еукаріот більше за обсягом, ніж у прокаріот (див. Розд. 3.6.3). Він розташований в основному в особливих ядерно них структурах - хромосомах (Див. Розд. 3.5.2), які відділені від цитоплазми ядерною оболонкою. Необхідний для синтезу білків апарат, що складається з рибосом, тРНК, набору амінокислот і ферментів, знаходиться в цитоплазмі клітини.
Значні відмінності є в молекулярної організації генів еукаріотичної клітини. У більшості з них кодують послідовності екзони перериваються ІНТРОН ділянками, які не використовуються при синтезі тРНК, рРНК або пептидів. Кількість таких ділянок варіюється в різних генах. Встановлено, що ген овальбуміна курей включає 7 интронов, а ген проколлагена ссавців - 50. Ці ділянки видаляються з первинно-транскрі- біруемой РНК, в зв'язку з чим використання генетичної інформації в еукаріотичної клітці відбувається дещо інакше. У прокариотичної клітці, де спадковий матеріал і апарат біосинтезу білка просторово НЕ роз'єднані, транскрипція і трансляція відбуваються майже одночасно. У еукаріотичної клітці ці два етапи не тільки просторово відокремлені ядерною оболонкою, але і в часі їх розділяють процеси дозрівання мРНК, з якої повинні бути видалені неінформативні послідовності (рис. 3.35).
Крім зазначених відмінностей на кожному етапі експресії генетичної інформації можна відзначити деякі особливості перебігу цих процесів у про- і еукаріот.
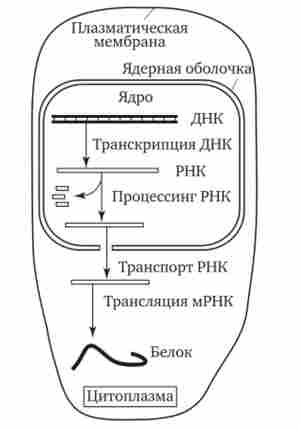
Мал. 3.35. Узагальнена схема процесу експресії генетичної інформації в еукаріотичної клітці
Транскрипція у про- і еукаріот. Транскрипція - це синтез РНК на матриці ДНК. У прокаріотів синтез усіх трьох видів РНК каталізується одним складним білковим комплексом -
РНК-полімераза.
Транскрипційний апарат еукаріотів включає три ядерні РНК-полімерази, а також РНК-полімерази мітохондрій і пластид. РНК-полімераза I виявляється в ядерцях клітин і відповідає за транскрипцію генів рРНК. РНК-полімераза II локалізується в ядерному соку і відповідає за синтез попередника мРНК. РНК-полімераза III - невелика фракція, яка перебуває в ядерному соку і здійснює синтез малих рРНК і тРНК. Кожен з цих ферментів має дві великі субодиниці і до 10 малих. РНК-полімерази мітохондрій і пластид відрізняються від ядерних.
Ферментний комплекс РНК-полімерази специфічно дізнається якусь нуклеотидну послідовність (часто не одну), розташовану на певній відстані від стартової точки транскрипції, - промотор. Стартовою точкою вважають нуклеотид ДНК, якому відповідав би перший нуклеотид, що включається ферментом в РНК-транскрипт.
У прокаріотів зазвичай недалеко від стартової точки проти ходу транскрипції розташовується послідовність з шести нуклеотидів - ТАТААТ, звана блоком Прібнова. Це середньостатистична послідовність, що складається з найбільш часто зустрічаються підстав, найконсервативнішими з яких є 1, 2 і 6-е підстави. Наявність в цій послідовності підстав, переважно з'єднаних подвійними водневими зв'язками з комплементарними основами іншого ланцюжка, очевидно, полегшує локальне плавлення подвійної спіралі ДНК і утворення двох її одноланцюгових ділянок при контакті з РНК-полімерази. Блок Прібнова розташовується в положенні від -11 до -5 або від -14 до -8, т. Е. За кілька нуклеотидів перед стартовою точкою транскрипції
(Рис. 3.36). Виявляючи цю послідовність, РНК-полімераза міцно зв'язується з нею і починає синтез РНК.
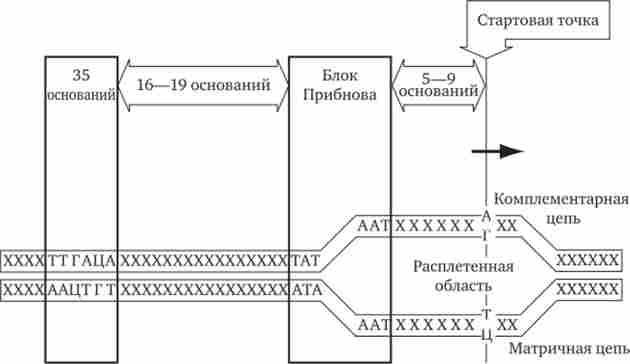
Мал. 3.36. Точки контакту для РНК-полімерази, що знаходяться у верхній ланцюга ДНК (промотор)
Настільки ж важлива роль у встановленні контакту РНК-полімерази з ДНК належить іншій нуклеотидної послідовності, центр якої знаходиться в положенні -35. її називають областю впізнавання - ТТГАЦА. Між двома зазначеними ділянками відстань досить постійно і становить від 16 до 19 пар нуклеотидів (п. Н.).
Промотори еукаріотичних генів також включають щонайменше дві специфічні нуклеотидні послідовності, центри яких знаходяться в положенні -25 і -75 п. Н. На відстані
19-27 нуклеотидів від стартової точки проти ходу транскрипції у багатьох генів еукаріот виявлена середньостатистична послідовність тата Ау (ТАТА-блок, або блок Хогнесса), в якій, так само як в блоці Прібнова у прокаріотів, переважають підстави, що утворюють більш слабкі зв'язки.
Другу послідовність, зустрічається в багатьох промоторах еукаріот і складається з ГГ ^ ЦААТЦТ, позначають як ЦААТ-блок. Вона займає положення між -70 і -80 нуклеотидами і також є областю, впізнаваною полимеразой.
У деяких генах виявлені багатокомпонентні промотори. Так, в окремих генах вірусу герпесу для ефективної ініціації транскрипції необхідні три послідовності ДНК, розташовані між -19 і -27, між -47 і -61, а також між -80 і -105 нуклеотидами.
Особливості промоторних дільниць свідчать про те, що для ініціації транскрипції має значення не тільки поєднання підстав в певних областях промотора, але і взаємне розташування в молекулі ДНК цих областей, з якими зв'язується ферментний комплекс РНК-полімерази.
Після встановлення контакту між РНК-полімерази і про- моторним ділянкою починається збірка молекули РНК, в яку першим найчастіше включається нуклеотид, що несе пуриновое підставу (як правило, аденін) і містить три 5'-фосфат них залишку. Далі, у міру просування РНК-полімерази уздовж молекули ДНК відбувається поступове подовження ланцюга РНК, яке триває до зустрічі ферменту з областю термінатора (див. Розд. 3.4.3.1). Термінатор - це ділянка, де припиняється подальше зростання ланцюга РНК і відбувається її звільнення від матриці ДНК. РНК-полімераза також відділяється від ДНК, яка відновлює свою двухцепочечную структуру.
У прокаріотів клітинах термінатори обов'язково містять паліндроми - дволанцюжкові послідовності нуклеотидів ДНК, які однаково читаються в обох напрямках (рис. 3.37). Ділянка РНК, транскрибуватися з такій послідовності, здатний утворювати дволанцюжкові шпильки за рахунок комплементарного спаровування нуклеотидів паліндрома. Можливо, це і є сигналом для завершення транскрипції, впізнаваним РНК-полімерази (рис. 3.38). Виникаючі шпильки, мабуть, зупиняють полімеразу на термінаторі. Слідом за шпилькою в молекулу РНК включається послідовність з нуклеотидів, що містять урацил (поліуретан), яка, ймовірно, бере участь у вивільненні РНК від матриці ДНК. Дійсно, поліуретан-послідовність РНК, поєднана з поліаденіловой (Поліани) послідовністю ДНК, характеризується слабкою взаємодією. Звертає на себе увагу той факт, що ділянка ДНК, багатий парами А-Т, зустрічається не тільки в місці ініціації транскрипції (блок Прібнова), але і в термінаторной області.
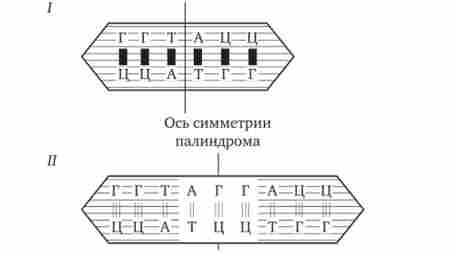
Мал. 3.37. Область ДНК з подвійною симетрією - паліндром.
I - паліндром, в якому є послідовність, однакова при читанні в протилежних напрямках; II - паліндром, в якому заштрихований інвертований повтор знаходиться на відстані від осі симетрії
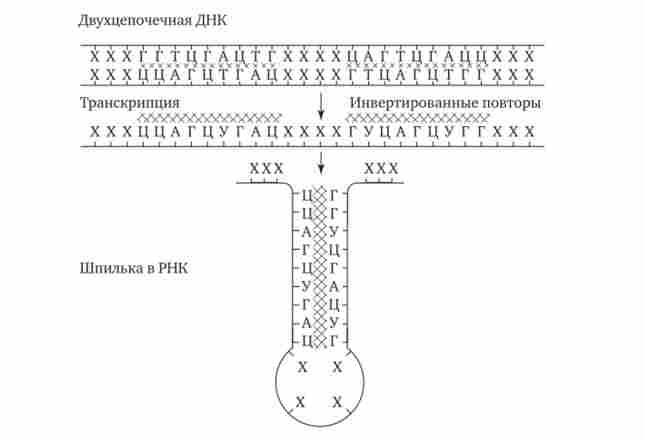
Мал. 3.38. Освіта шпильки ділянкою РНК при термінації транскрипції у прокаріот.
Область РНК, несуча паліндром, утворює комплементарно спаровуються структуру - шпильку (інвертовані повтори заштриховані)
Бактеріальні термінатори значно різняться за своєю ефективністю. Деякі з них як би не помічаються РНК-полімерази, і вона продовжує транскрипцію за межами термінатора. Таке просочування термінатора при транскрипції бактеріальних генів спостерігається в результаті запобігання термінації специфічними білками - факторами антітермінацш. Наслідком антітермінаціі є синтез поліцістронной мРНК, що включає в себе інформацію, списану з декількох послідовно розташованих структурних генів.
Термінатори еукаріотичних генів вивчені в меншій мірі, ніж у проскаріот, але в них також виявлені райони, багаті Г-Ц парами, з'єднаними потрійними водневими зв'язками, в яких розташовується ділянку з А-Т парами. На цій ділянці в транскрипт включається поліуретан-послідовність, слабо взаємодіє з матричної Поліани-областю ДНК.
Можливо, область термінатора, багата Г-Ц парами, грає певну роль в зупинці РНК-полімерази, а ділянку РНК, що містить yySv, забезпечує відділення транскрипту від матриці ДНК.
У еукаріот не виявлено освіти структур, подібних шпильок в прокариотичних РНК. Тому, яким чином у них здійснюється термінація транскрипції, залишається неясним.
У складі всіх мРНК можна виділити кодують ділянки, що представляють набір кодонів, які шифрують послідовність амінокислот в пептиді. Як правило, ці ділянки починаються стартовим кодоном АУТ, але іноді у бактерій використовується кодон ГУТ. На кінці кодує розташовується терминирующего кодон. Крім кодують ділянок в мРНК на обох кінцях можуть розташовуватися додаткові послідовності. На 5'-кінці це лідерних ділянку, розташовану перед стартовим кодоном. На -кінці - трейлер, наступний за кодоном-термінатором.
У поліцістронной мРНК прокаріотів між кодують ділянками є межцістронние області, що варіюються за розмірами (рис. 3.39).
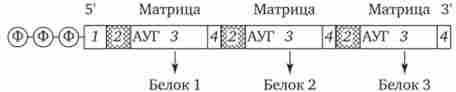
Мал. 3.39. Поліцістронной матрична РНК прокариот.
1 - некодуючі області, 2 - межцістронние області, 3 - кодують області, 4 - терминирующего кодони
У зв'язку з тим що прокариотичні гени цілком складаються з нук- леотідних послідовностей, що беруть участь в кодуванні інформації, транскрибоване з них РНК відразу після їх синтезу здатні виконувати функцію матриць для трансляції. Лише у виняткових випадках потрібно їх попереднє дозрівання - процесинг.
На відміну від прокаріотів генів більшість генів еукаріотів переривчасті, так як несуть у своєму складі неінформативні нуклеотидні послідовності - інтрони, які беруть в кодуванні інформації. У зв'язку з цим первинні транскрипти, синтезовані РНК-полімераза II, мають більші, ніж необхідно для трансляції, розмірами і виявляються менш стабільними. В сукупності вони утворюють так звану гетерогенную ядерну РНК (гяРНК), яка, перш ніж вийти з ядра і почати активно функціонувати в цитоплазмі, піддається процесингу і перетворюється в зрілі мРНК.
Дозрівання, або процесинг, мРНК передбачає зміну первинного транскрипту і видалення з нього не кодують інтронних ділянок з подальшим з'єднанням (сплайсингом) Кодують послідовностей - екзонів. Модифікування первинного транскрипту еукаріотичної мРНК починається незабаром після синтезу його 5'-кінця, що містить одне з пуринових підстав (аденін або гуанін). На цьому кінці утворюється ковпачок - кеп, який блокує
5'-кінець мРНК шляхом приєднання до першого нуклеотиду транскрипта тріфосфонуклеозіда, що містить гуанін, зв'язком 5'-5 '
В результаті утворюється послідовність ГфффАфИ ..., в якій залишок гуаніну знаходиться в зворотній орієнтації по відношенню до інших нуклеотидам мРНК. Модифікація 5'-кінця мРНК передбачає також метилювання приєднаного гуаніну та перших двох-трьох підстав первинного транскрипту (рис. 3.40). Утворені на 5'-кінцях мРНК кепи забезпечують впізнавання молекул мРНК малими субчастиц рибосом в цитоплазмі. Копіювання здійснюється ще до закінчення синтезу первинного транскрипту.
Після завершення транскрипції відбувається видалення частини нуклеотидів на -кінці первинного транскрипту і приєднання до нього послідовності, що складається з 100-200 залишків аде- Нілов кислоти (Поліани) (див. Рис. 3.40). Вважають, що ця послідовність сприяє подальшому процесингу і транспорту зрілої мРНК з ядра. Після виходу мРНК в цитоплазму її Поліани-послідовність поступово коротшає під дією ферментів, відщеплюється нуклеотиди на -кінці. Таким чином, по довжині Поліани-послідовності можна побічно судити про час перебування мРНК в цитоплазмі. Можливо, додавання Поліани-послідовності в ході процесингу підвищує стабільність мРНК. Однак близько третини мРНК взагалі не містять Поліани-ділянки. До них відносяться, наприклад, гістонові мРНК.
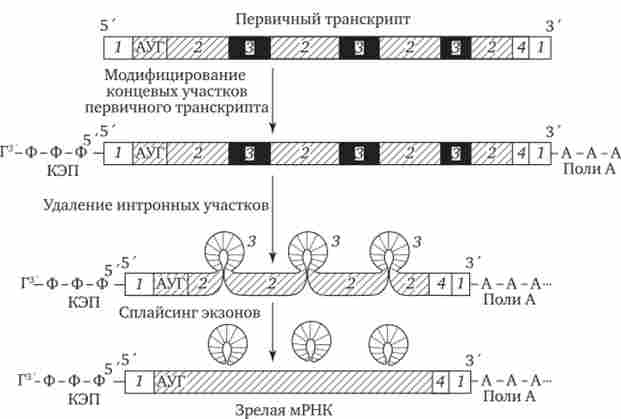
Мал. 3.40. Освіта зрілої мРНК еукаріотів в ході процесингу.
1 - некодуючі послідовності, 2 - екзонів, 3 - інтрони, 4 - кодон-термінатор
Освіта кепа на 5'-кінці і Поліани-послідовності на -кінці характерно тільки для процесингу РНК, що синтезуються РНК-полімерази І. Крім метилування при формуванні кепів в мРНК вищих еукаріот відбувається метилування невеликої частини внутрішніх нуклеотидів з частотою приблизно один на тисячу підстав мРНК.
Поряд з модифицированием мРНК еукаріотів процесинг припускає видалення з первинних транскриптів неінформативних для даного білка інтрони ділянок, розмір яких варіює від 100 до 10 000 нуклеотидів і більше. На частку інтронів припадає близько 80% всієї гяРНК. Видалення інтронів з подальшим з'єднанням екзонних ділянок називають сплайсингом (Див. Рис. 3.40).
Сплайсинг є механізм, який повинен забезпечувати видалення з первинного транскрипту суворо визначених інтрони ділянок. Порушення цього процесу може привести до зсуву рамки зчитування при трансляції та неможливості синтезу нормального пептиду. Закономірність вирізання інтронів, очевидно, забезпечується завдяки наявності на їх кінцях специфічних нуклеотидних послідовностей, службовців сигналами для сплайсингу.
В даний час описано кілька ймовірних механізмів сплайсингу, що забезпечують точність цього процесу. Можливо, вона досягається дією якихось ферментів, специфічно котрі дізнаються кінцеві ділянки интронов і каталізують розрив фосфо діефірних зв'язків на кордоні екзон - інтрон, а потім утворення зв'язків між двома екзонами.
Встановлено активну участь в сплайсинге особливих малих, ядерно них РНК (мяРНК), що утворюють комплекси з білками (мяРНП). Очевидно, мяРНК своїми нуклеотидними послідовностями комплементарно взаємодіють з кінцевими ділянками інтронів, які утворюють при цьому замкнуті петлі. Розщеплення РНК в гирлі інтронів петлі призводить до видалення неінформативної послідовності і з'єднанню (сплайсингу) зближених решт екзонів.
Обговорюється також автокаталітічна здатність РНК-тран- скрипта до сплайсингу. Описані способи сплайсингу свідчать про відсутність універсального механізму цього процесу, проте у всіх випадках досягається точне видалення інтронів з утворенням певної мРНК, що забезпечує синтез необхідного клітці білка.
В даний час доведено можливість альтернативного (взаємовиключення) сплайсингу, при якому з одного і того ж первинного транскрипту можуть віддалятися різні нуклеотидні послідовності і утворюватися різні зрілі мРНК. В результаті одна і та ж послідовність нуклеотидів ДНК може служити інформацією для синтезу різних пептидів. Альтернативний сплайсинг, ймовірно, дуже характерний в системі генів імуноглобулінів у ссавців, де він дозволяє формувати на основі одного транскрипту мРНК для синтезу різних видів антитіл.
Завдяки перетворенням, що відбувається з РНК-транскриптом в ході процесингу, зрілі мРНК еукаріотів характеризуються більшою стабільністю в порівнянні з прокариотичними мРНК.
По завершенні процесингу зріла мРНК проходить відбір перед виходом в цитоплазму, куди потрапляє всього 5% гяРНК. Інша частина розщеплюється, не залишаючи ядра.
Таким чином, перетворення первинних транскриптів еукаріотичних генів, зумовлені їх екзон-ітронной організацією і необхідністю переходу мРНК з ядра в цитоплазму, визначають особливості реалізації генетичної інформації в еукаріотичної клітці.
У прокаріотів клітинах процес трансляції пов'язаний з синтезом мРНК: вони відбуваються практично одночасно. В значній мірі це пов'язано з недовговічністю бактеріальної мРНК, яка досить швидко піддається розпаду. Взаємопов'язаність транскрипції і трансляції у бактерій проявляється в узгодженості швидкостей цих процесів. При 37 ° С транскрипція йде зі швидкістю 2500 нуклеотидів / хв (14 кодонів / с), а трансляція здійснюється зі швидкістю 15 амінокислот / с.
Трансляція у прокаріот починається незабаром після утворення 5'-кінця мРНК, раніше, ніж закінчується її синтез. В результаті слідом за РНК-полімерази по мРНК рухаються рибосоми, здійснюють збірку пептидних ланцюгів (рис. 3.41). Через деякий час після початку транскрипції (близько 1 хв) і до завершення трансляції -кінця матриці починається деградація її 5'-кінця. З огляду на те що час життя різних мРНК не однаково, кількість білка, синтезованого на різних матрицях, по-різному.
Однією з особливостей трансляції у прокаріотів є включення в пептидную ланцюг в якості першої амінокислоти модифікованого метіоніну - формілметіонін, з якого починаються всі знову синтезовані пептиди. Навіть в тому випадку, коли роль стартового кодону виконує кодом ГУТ, в звичайних умовах шифрує валін, в першому положенні пептиду виявляється формілметі- Онін. Стартовий кодон АУТ або ГУТ слід за лідерних ділянкою, який екранується рибосомою в момент ініціації трансляції.
З'єднання рибосоми з мРНК обумовлено комплементарною взаємодією нуклеотидів однієї з рРНК з нуклеотидної послідовністю лідера мРНК.
Ця послідовність (Шайна - Дальгарно) розташовується на відстані 4-7 підстав перед кодоном АУГ і виявляється повсюдно в лідерних ділянках у прокаріот.
При з'єднанні 5'-кінця мРНК з малої субчастиц рибосоми стартовий кодон зазвичай виявляється майже в середині екранованого рибосомою фрагмента мРНК, в області, що відповідає її П-ділянці.
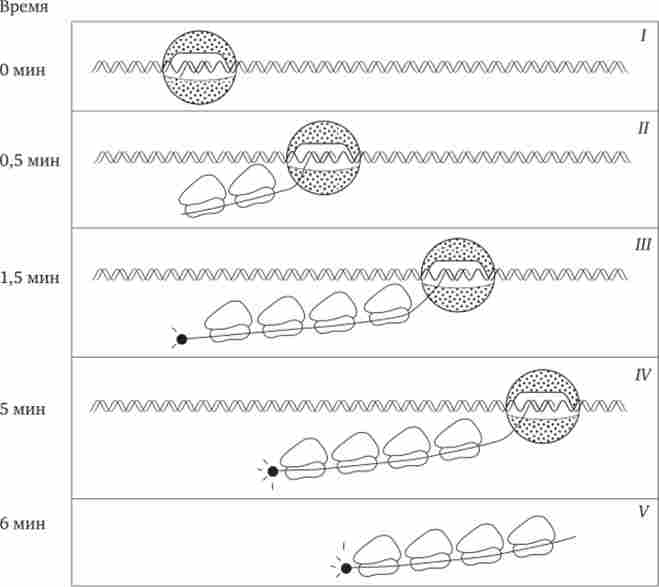
Мал. 3.41. Транскрипція, трансляція і деградація мРНК у прокаріот.
/ - РНК-полімераза зв'язується з ДНК і починає синтезувати мРНК в напрямку 57 -> З7; II - у міру просування РНК-полімерази до 57-кінця мРНК прикріплюються рибосоми, початківці синтез білка; III - група рибосом слід за РНК-полімерази, на 57-Наприкінці мРНК починається її деградація; IV - процес деградації протікає повільніше, ніж транскрипція і трансляція; V - після закінчення транскрипції мРНК звільняється від ДНК, на ній триває трансляція і деградація на 57-наприкінці
У еукаріот трансляція здійснюється в цитоплазмі, куди потрапляє з ядра зріла мРНК. Копіювати кінець мРНК розпізнається малою субчастиц рибосоми, потім лідируюча послідовність, що містить до 100 нуклеотидів, взаємодіє з рРНК. При цьому стартовий кодон АУТ виявляється в недобудованому П-ділянці рибосоми. Після приєднання до стартового кодону аміноацил-тРНК, що несе метіонін, відбувається возз'єднання двох субчастиц рибосоми і формуються її А- і П-ділянки. Синтез білка в еукаріотичної клітці, здійснюваний на моноцістронной мРНК, завершується після проходження рибосомою по всій мРНК, аж до впізнавання нею кодону-термінатора, який припиняє утворення пептидних зв'язків.
Синтезовані в ході трансляції пептидні ланцюга на основі своєї первинної структури набувають вторинну і третинну, а багато - і четвертинних організацію, утворену кількома пептидними ланцюгами. Залежно від функцій, які виконуються білками, їх амінокислотні послідовності можуть зазнавати різні перетворення, формуючи функціонально активні молекули білка.
Багато мембранні білки синтезуються у вигляді пре-білків, що мають на N-кінці лідерних послідовність, яка забезпечує їм впізнавання мембрани. Ця послідовність відщеплюється при дозріванні і вбудовуванні білка в мембрану. Секреторні білки також мають на N-кінці лідерних послідовність, яка забезпечує їх транспорт через мембрану.
Деякі білки відразу після трансляції несуть додаткові амінокислотні про-послідовності, що визначають стабільність попередників активних білків. При дозріванні білка вони видаляються, забезпечуючи перехід неактивного пробелкой в активний білок. Наприклад, інсулін спочатку синтезується як пре-про- інсулін. Під час секреції пре-послідовність відщеплюється, а потім проінсулін піддається модифікації, при якій з нього видаляється частина ланцюга і він перетворюється в зрілий інсулін.
Формуючи третинну і четвертинних організацію в ході посттрансляційних перетворень, білки набувають здатність активно функціонувати, включаючись в певні клітинні структури і здійснюючи ферментативні і інші функції.
Розглянуті особливості реалізації генетичної інформації в про- і клітині виявляють принципову подібність цих процесів. Отже, механізм експресії генів, пов'язаний з транскрипцією і наступною трансляцією інформації, яка зашифрована за допомогою біологічного коду, склався в цілому ще до того, як були сформовані ці два типи клітинної організації. Дивергентна еволюція геномів про- і еукаріот привела до виникнення відмінностей в організації їх спадкового матеріалу, що не могло не відбитися і на механізмах його експресії.
Постійне вдосконалення наших знань про організацію та функціонування матеріалу спадковості і мінливості обумовлює еволюцію уявлень про ген як функціональну одиницю цього матеріалу.