Активний (т. Е. Енергозалежний) транспорт молекул через мембрану проти градієнта концентрації здійснюється за участю мембранних білків, які використовують для процесу транслокації енергію гідролізу АТФ. На відміну від пасивного транспорту, який йде мимоволі, білки-переносники повинні не тільки транспортувати молекулу через мембрану, а й володіти АТФ-азним дією, т. Е. Каталізувати гідроліз АТФ, який є основним джерелом енергії для активного транспорту. Залежно від способу використання енергії для транспорту молекул виділяють первинно і вторинно-активний транспорт.
при первинно-активному транспорті донором енергії є безпосередньо молекула АТФ і процес перенесення речовини через мембрану супроводжується її гідролізом.
при вторинно активному транспорті градієнт іонів (Na+ До+, Н+ і лр.), створений на мембрані функціонуванням систем первинно-активного транспорту, використовується для транспорту інших молекул, наприклад вуглеводів, деяких амінокислот, аніонів та ін.
Відомі три основних типи первинно-активного транспорту іонів:
натрій-калісвий насос - №+/ К+-аденозинтрифосфатаза (Иа+/ К+-АТФ-аза), який переносить іони натрію з клітини, а калію - в клітку;
кальцієвий насос - Са2+-АТФ-аза, який транспортує Са2+ з клітки або цитозоля в саркоплазматичний ретикулум;
Н+-АТФ-аза - протонний насос, що функціонує в сполучають мембранах, в тому числі в мітохондріальної мембрани, де Н+-АТФ-аза працює в зворотному напрямку, використовуючи ДРН+, утворюється вдихательная ланцюга для синтезу АТФ (гл. 15).
Для активного транспорту, як і для полегшеної дифузії, характерні висока специфічність, ефект насичення транспортних білків транспортуються молекулами, коли кінетична крива виходить на плато, а також дію інгібіторів.
Як приклад первинно-активного транспорту можна привести транспорт, який здійснюється №+/ К+-АТФ-азой, як однієї з найбільш важливих і широко поширених активних транспортних систем в плазматичній мембрані тварин клітин. Ця система, що отримала назву Na+-K * -Hacoca, відповідає за підтримання в клітці високої концентрації К+ і низькою Na+ шляхом перенесення До+ всередину клітини, a Na+ з клітки назовні проти градієнта їх концентрації і тому вимагає витрати АТФ. Виявляється, в тваринній клітині внутрішньоклітинна концентрація іонів калію приблизно в 30 разів вище, а іонів натрію в 10 разів нижче, ніж у навколишньому середовищі. Така асиметрія іонного складу визначає вміст води і іонний склад в клітці, електричну збудливість нервових і м'язових волокон, служить рушійною силою для транспорту в клітку цукрів і амінокислот, є важливим фактором у процесі біосинтезу білка.
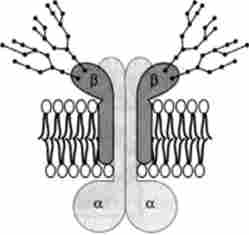
Мал. 22.8. структура Na+, К * -залежною АТФ-ази
№+/ К+-АТФ-аза була відкрита в 1957 р Й. Скоу у фракції плазматичних мембран нервів краба, згодом вона була виявлена у всіх досліджених клітинах тварин, особливо велике се зміст у органах, що здійснюють інтенсивний сольовий обмін (нирки) або виконують електричну роботу (мозок, нерви).
№+/ К+-АТФ-аза є олігомерного білок, що складається з субодиниць двох типів (а і Р), що входять до складу ферменту в еквімолярних кількостях. Велика а-субодиниця (-112 kDa) формує каталитично активний центр, який здійснює гідроліз АТФ; менша р-субодиниця (-45 kDa) глікозильованого, при цьому вуглеводні ланцюги експоновані на зовнішній стороні мембрани (рис. 22.7).
Активний фермент містить як мінімум 2а- і 2р-суб'сдініци, які щільно інтегровані в мембрану і активуються іонами Mg2+. В даний час експериментально доведено, що Иа+/ К+-АТФ-аза є як енергопреобразующей частиною «Na+/ K+-насоса », так і сама здійснює активний транспорт Na+ і К+, пов'язаний з гідролізом АТФ, т. е. переносить катіони проти градієнта концентрації. Описано два кон- формаційних стану АТФ-азного комплексу з різним енергетичним рівнем, які прийнято позначати?, І? ^. Конформація?, Має канал, відкритий всередину клітини, і ділянки, специфічно зв'язують іони
Na+ (Дсфосфорілірованная форма АТФ-ази). При переході в конформацію Е2 відбувається переорієнтація комплексу в мембрані, канал відкритий на зовнішню сторону мембрани і фермент специфічно зв'язує іони К+ (Фосфорілірованний форма АТФ-ази).
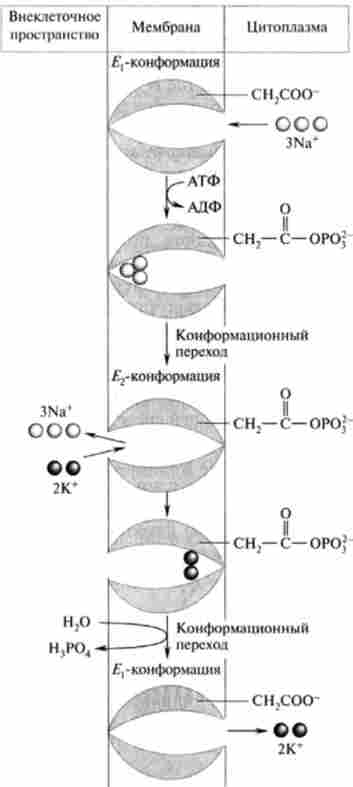
За загальноприйнятою поданням, механізм дії АТФ-ази включає кілька стадій (рис. 22.9).
Мал. 22.9. Механізм функціонування NaVK ^ -АТФ-ази
1. Молекула АТФ з цитоплазми зв'язується з активним центром однієї з а-суб'сдініц АТФ-ази (конформація?,). Приєднання АТФ супроводжується зв'язуванням трьох іонів натрію, що активує каталітичну активність ферменту, відбувається гідроліз АТФ, при цьому фосфорилируется карбоксильная група бічного ланцюга залишку аспарагінової кислоти інший субодиниці.
2. Фосфорилування ферменту призводить до переходу його в конформацію Е2і переорієнтації в мембрані таким чином, що пов'язані 3Na+ виводяться через канал, відкритий на зовнішню сторону, оскільки АТФ-аза втрачає спорідненість до цих іонів.
3. Конформація Е2 ферментного білка має сайти, специфічні до іонів К+, і два іона К+ приєднуються до іонсвязивающім центрам фосфорилированного білка.
4. Відбувається (можливо, мимовільний) гідроліз фосфоефір- ної зв'язку і дефосфорілірованіе ферменту, що викликає його перехід в вихідну конформацію? ,, при якій іонний канал відкритий всередину клітини, через нього два іона К+ виходять в цитоплазму.
На кожну витрачену молекулу АТФ фермент переносить три іона Na+ з клітки і два іона К+ в клітку, т. е. перенесення катіонів нееквівалентний. Таким чином, на мембрані виникає різниця концентрацій і електричних потенціалів, і третій іон К+ надходить в клітку вже за механізмом вторічноактівного транспорту.
Базальні ядра - анатомія центральної нервової системи Базальні ядра включають хвостате ядро, сочевицеподібне ядро, огорожу, мигдалеподібне тіло і прилегле ядро. Найбільшим з цих ядер є хвостате ядро ( п. caudatus). Воно витягнуте в ростри-каудальному напрямку (спереду назад) і має С-подібну форму (рис. 9.1). Мал. 9.1. Базальні ядра: пунктиром
Базальні ганглії - нервова система: анатомія, фізіологія, Нейрофармакологія До складу базальних гангліїв включають хвостате ядро, бліда куля, шкаралупу, огорожу і мигдалеподібне тіло (див. Рис. 7.2; рис. 7.6). Найбільшою з перерахованих структур є хвостате ядро. Це ядро витягнуто в ростри-каудальному напрямку і має С-подібну форму. Передня частина хвостатого ядра
Автономна (вегетативна) нервова система - анатомія центральної нервової системи В результаті вивчення даного розділу студент повинен: знати відмінні риси ВНС в порівнянні з іншими нейроанатомічсскімі і функціональними блоками мозку; принципи поділу ВНС на симпатичну, парасимпатичну і метасім- патичної складові; їх взаємодія; центральні і периферичні компоненти ВНС; їх
Аспекти вуглеводного обміну в відновної медицини - біохімія людини Актуальним завданням медицини є терапія патології метаболізму вуглеводів при одночасній патології ліпідного метаболізму. Це обумовлено різким зростанням числа діабетичних пацієнтів з ознаками ожиріння. Таким чином, регуляція концентрації глюкози в крові (глікемії) у хворих на цукровий діабет
Анестетики - стоматологія. Ендодонтія Ін'єкційна анестезія вимагає введення анестетика в тканини. Амідні анестетики представлені 0,25% -ним, 0,5% -ним, 1,0% -ним, 2,0% -ним розчинами лідокаїну або ксілокаіна, 2,0% -ним розчином тримекаина, 3,0% -ним розчином мепівакаіна або скандонеста, 4% -ним розчином артикаина (ультракаїн,
Аналізатори - цитологія, гістологія і ембріологія Аналізатори - чутливі відділи рефлекторних дуг, що забезпечують зв'язок центральної нервової системи з навколишнім середовищем. В аналізаторі розрізняють три частини: периферичну, або рецептори, що сприймають подразнення; проміжну - провідні шляхи і підкіркові освіти (передають імпульси);
Альтернативні теорії харчування - фізіологія харчування В останні десятиліття з'явилося багато нових оригінальних теорій харчування, які не вписуються в рамки традиційних уявлень про харчування, але в той же час мають глибоке історичне коріння. Безсумнівно, в кожній з цих теорій є логіка і раціональне зерно. Однак для вибору своєї методики харчування