| Головна |
| «« | ЗМІСТ | »» |
|---|
Сутність стадії гаструляції полягає в тому, що одношаровий зародок - бластула - перетворюється в багатошаровий - дво- або тришаровий, званий гаструлою (Від грец. гастер - шлунок в зменшувально сенсі).
У примітивних хордових, наприклад у ланцетника, однорідна одношарова бластодерма під час гаструляції перетворюється в зовнішній зародковий листок - ектодерму - і внутрішній зародковий листок - ентодерму. Ентодерма формує первинну кишку з порожниною всередині - гастроцель. Отвір, що веде в гастроцель, називають бластопором або первинним ротом. Два зародкових листка є визначальними морфологічними ознаками гаструляції. Їх існування на певній стадії розвитку у всіх багатоклітинних тварин, починаючи з кишковопорожнинних і закінчуючи вищими хребетними, дозволяє думати про гомології зародкових листків і єдності походження всіх цих тварин.
У хребетних крім двох згаданих під час гаструляції утворюється ще третій зародковий листок - мезодерма, що займає місце між екто- і ентодерми. Розвиток середнього зародкового листка, що представляє собою хордомезодерми, є еволюційним ускладненням фази гаструляції у хребетних і пов'язане з прискоренням у них розвитку на ранніх стадіях ембріогенезу. У більш примітивних хордових тварин, таких як ланцетник, хор- домезодерма зазвичай утворюється на початку наступного після гаструляції фази - органогенезу. Зсув часу розвитку одних органів щодо інших у нащадків в порівнянні з предкової групи є проявом гетерохронии. Зміна часу закладки найважливіших органів в процесі еволюції зустрічається нерідко.
Процес гаструляції характеризується важливими клітинними перетвореннями, такими як спрямовані переміщення груп і окремих клітин, виборче розмноження і сортування клітин, початок цітодіфференціровкі і індукційних взаємодій. Перераховані клітинні механізми онтогенезу детально розбираються в розд. 8.2.
Способи гаструляції різні. Виділяють чотири різновиди спрямованих в просторі переміщень клітин, що призводять до перетворення зародка з одношарового в багатошаровий.
інвагінація - впячивание одного з ділянок бластодерми всередину цілим пластом. У ланцетника впячиваются клітини вегетативного полюса, у земноводних інвагінація відбувається на кордоні між анімальний і вегетативним полюсами в області сірого серпа. Процес інвагінації можливий тільки в яйцях з невеликою або середньою кількістю жовтка.
епіболія - обростання дрібними клітинами анімального полюса більших, які відстають у швидкості ділення і менш рухливих клітин вегетативного полюса. Такий процес яскраво виражений у земноводних.
деламінація - розшарування клітин бластодерми на два шари, що лежать один над одним. Деламінація можна спостерігати в дискобластула зародків з частковим типом дроблення, таких як плазуни, птахи, яйцекладущие ссавці. Деламінація проявляється в ембріобласта плацентарних ссавців, приводячи до утворення гіпобласта і епібласта.
імміграція - переміщення груп або окремих клітин, не об'єднаних в єдиний пласт. Імміграція зустрічається у всіх зародків, але найбільшою мірою характерна для другої фази гастру- ляции вищих хребетних.
У кожному конкретному випадку ембріогенезу, як правило, поєднуються кілька способів гаструляції.
Більш детальний розгляд гаструляції у ланцетника, жаби, курчати і ссавців, до чого ми переходимо, допоможе глибше зрозуміти еволюційні зв'язку і розібратися в закономірностях індивідуального розвитку.
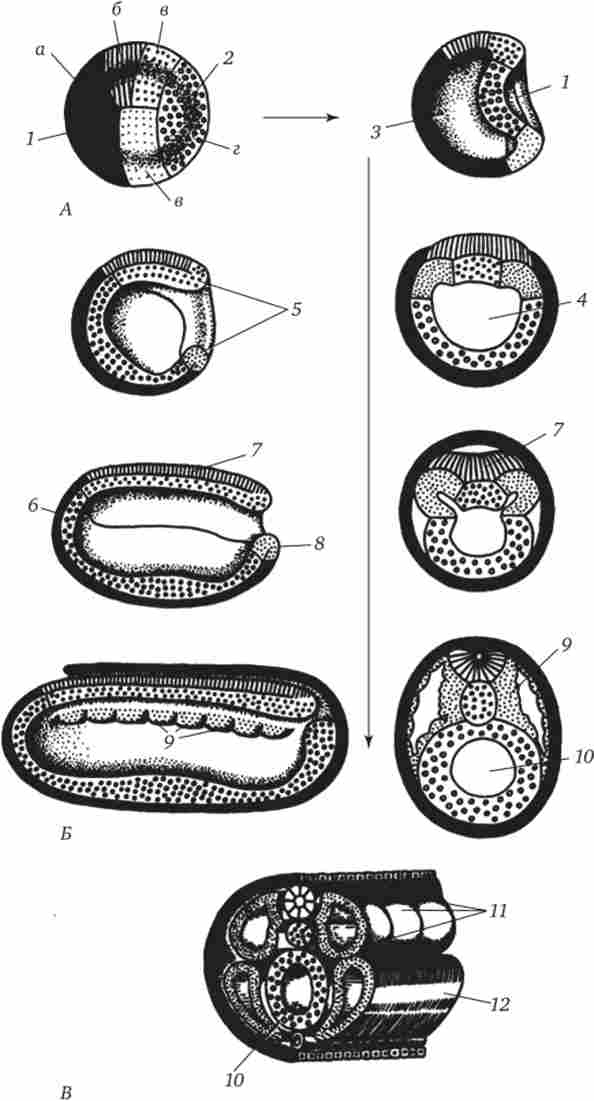
гаструляция ланцетника показана на рис. 6.3. Різними маркерами на стадії бластули (рис. 6.3, А) відзначені презумптівного зачатки. Це області бластули, з клітинного матеріалу яких в ході гаструляції і раннього органогенезу (нейруляции) зазвичай утворюються цілком певні зародкові листки і органи (рис. 6.3, Б і В).
Інвагінація починається на вегетативному полюсі. Через більш швидкого ділення клітини анімального полюса розростаються і штовхають всередину бластули клітини вегетативного полюса. Цьому сприяє зміна стану цитоплазми в клітинах, що утворюють губи бластопора і прилеглих до них. Внаслідок инвагинации бластоцель зменшується, а гастроцель збільшується. Одночасно зі зникненням бластоцель ектодерма і ентодерми приходять в тісний контакт. У ланцетника, як і у всіх вторичноротих тварин (до них відносять тип Иглокожие, тип Хордові і деякі інші нечисленні типи тварин), область бластопора перетворюється в хвостову частину організму на відміну від первичноротих, у яких бластопор відповідає головній частині. Ротовий отвір у вторичноротих утворюється на протилежному бла стопору кінці зародка.
Гаструляция у земноводних має багато спільного з гаструляціей ланцетника, але так як в яйцеклітинах у них жовтка набагато більше і розташований він переважно на вегетативному полюсі, великі бластомери амфібластули не здатні впячиваются всередину. інвагінація проходить трохи інакше. На кордоні між ані- ною і вегетативним полюсами в області сірого серпа клітини спочатку сильно витягуються всередину, набуваючи вигляду колбовідних
I - анімаль- ний полюс,
Мал. 6.3. Презумптівного зачатки, гаструляция і нейруляція у ланцетника.
і черевна губи бластопора,
II - сегментовані соміти,
12 - черевна частина мезодерми; а, б, в, г, д - позначення презумптів- них і розвиваються органів:
а - ектодерма шкірна,
6 - нервова трубка, в - хорда, г - Ендотерм, епітелій кишки,
д - мезодерма
Л - презумптівного зачатки на стадії бластули (вид зовні) і ранньої гаструли (вид на зрізі); Б - пізня гаструла і нейруляція на сагиттальном (лівий ряд) і поперечному (правий ряд) розрізах; В - пластична модель зародка в кінці періоду нейруляции
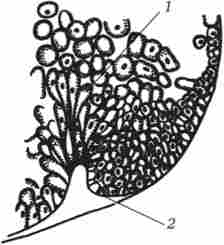
Мал. 6.4. Колбовідние клітини в області бластопора ранньої гаструли амфібій.
Одночасно більш дрібні клітини анімального полюса, діляться швидше, починають переміщатися в бік вегетативного полюса. В області спинний губи вони підвертаються і впячиваются, а з боків і з боку, протилежного серповидної бороздке, покривають собою більші клітини вегетативного полюса. потім процес епіболії призводить до того, що утворюються бічні і черевна губи бластопора. Бластопор змикається в кільце, всередині якого деякий час видно великі світлі клітини вегетативного полюса у вигляді так званої желточной пробки. Пізніше вони повністю занурюються всередину, а бластопор звужується.
За допомогою методу маркування прижиттєвими (вітальними) барвниками у земноводних детально вивчені переміщення клітин бластули під час гаструляції. Встановлено, що конкретні області бластодерми, звані презумптівного зачатками (Від лат. Ргае- sumptio - припущення), при нормальному розвитку виявляються спочатку в складі певних зачатків органів, а потім в складі самих органів (рис. 6.5). Відомо, що у безхвостих амфібій матеріал презумптівного хорди і мезодерми на стадії бластули лежить не на її поверхні, а у внутрішніх шарах стінки амфібластули, проте приблизно на тих рівнях, як це показано на малюнку. Аналіз ранніх етапів розвитку земноводних дозволяє зробити висновок про те, що овоплазматічна сегрегація, яка чітко проявляється в яйцеклітині і зиготі (рис. 6.6), має велике значення у визначенні долі клітин, що успадкували ту чи іншу ділянку цитоплазми. Певну схожість процесів гаструляції і областей презумптівного органів у земноводних і ланцетника, т. Е. Гомология основних органів, таких як нервова трубка, хорда, вторинна кишка, вказує на їх філіпченкове спорідненість.
Гаструляция у зародків з меробластичні типом дроблення і розвитку має свої особливості. У птахів вона починається слідом за дробленням і утворенням бластули під час проходження зародка по яйцепроводу. До моменту відкладання яйця зародок вже складається з декількох шарів: верхній шар називають епібла- стом, нижній - первинним гіпобласт (Див. Рис. 6.2, В). Між ними знаходиться вузька щілина - бластоцель. потім утворюється вторинний гіпобласт, спосіб утворення якого не цілком ясний.
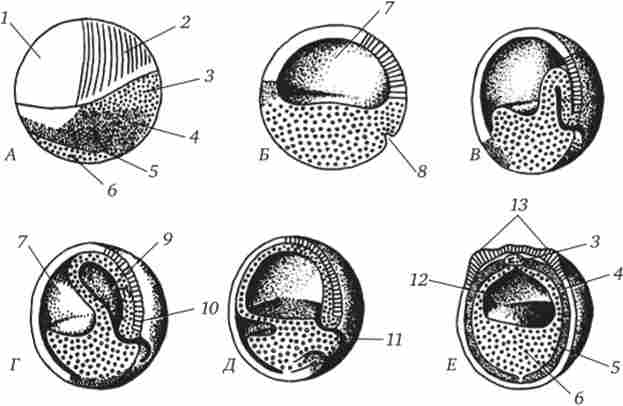
Мал. 6.5. Карта областей презумптівного зачатків органів на ранніх стадіях ембріонального розвитку амфібій.
А - стадія бластули (вид зліва); Б - 4 - послідовні етапи гаструляції (саггитальний зрізи); Е - початок нейруляции (поперечний зріз).
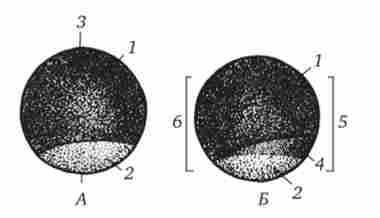
Мал. 6.6. Овоплазматічна сегрегація в ікринці трав'яний жаби.
ного по швидкості ділення клітин і переміщення їх з бічних ділянок епібласта до центру, назустріч один одному. Утворюється так звана первинна смужка, яка витягується в напрямку до головного кінця. У центрі первинної смужки утворюється первинна борозенка, а по краях - первинні
Є дані про те, що в первинному гіпобласт птахів беруть початок первинні статеві клітини, а вторинний - утворює внезародишевую ентодерму. Освіта первинного і вторинного гіпобласта розглядають як явище, що передує гаструляції.
Основні події гаструляції і остаточне утворення трьох зародкових листків починаються після відкладання яєць з початком інкубації. Виникає скупчення клітин в задній частині епібла- ста як результат неравномер-
А - відразу після запліднення;
Б - через 2 години після запліднення (вид зліва).
1 - пигментированная анімальна область, 2 - непігментованими негативна область, 3 - головно-хвостова вісь майбутнього організму, 4 - сірий серп, 5 - спинна сторона, 6 - черевна сторона валики. На головному кінці первинної смужки виникає потовщення - гензеновскій вузлик, а в ньому - первинна ямка (рис. 6.7).
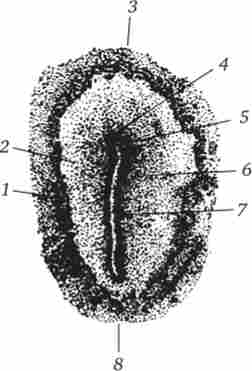
Мал. 6.7. Курячий зародок на стадії первинної смужки (вигляд із спинного боку).
1 - темна область, 2 - просвічує область зародкового диска, 3 - головний кінець, 4 - гензеновскій вузлик, 5 - первинна ямка, 6 - первинна борозенка, 7-первинний валик, 8 - хвостовий кінець
Коли клітини епібласта входять в первинну борозенку, їх форма змінюється. Вони нагадують за формою колбовідние клітини гаструли земноводних. Потім ці клітини набувають зірчасті форму і занурюються під епі- бласт, утворюючи мезодерму (рис. 6.8). Ентодерма утворюється на основі первинного і вторинного гіпобласта з додаванням нового покоління ентодермаль- них клітин, що мігрують з верхніх шарів бластодерми. Наявність декількох генерацій ентодермальних клітин вказує на розтягнутість періоду гаструляції в часі.
Частина клітин, мігруюча з епібласта через гензеновскій вузлик, утворює майбутню хорду. Одночасно із закладкою і подовженням хорди гензеновскій вузлик і первинна смужка поступово зникають в напрямку від головного до хвостового кінця. Це відповідає звуження і закриття бластопора. У міру скорочення первинна смужка залишає за собою сформовані ділянки осьових органів зародка в напрямку від головних до хвостовим відділам.
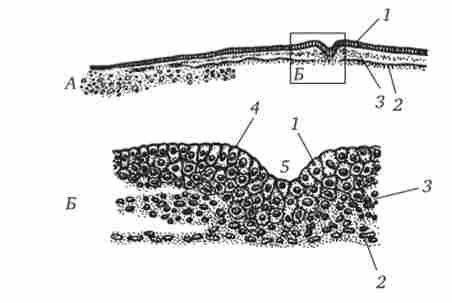
Мал. 6.8. Курячий зародок на стадії первинної смужки (поперечний зріз).
Л, Б - при малому і великому збільшенні;
1 - ектодерма, 2 - ентодерми, 3 - мезодерма, 4 - первинний валик, 5 - первинна борозенка
Видається обгрунтованим розглядати переміщення клітин в курячому зародку як гомологічні епіболії, а первинну смужку і гензеновскій вузлик - як гомологічні бластопора в спинний губі гаструли земноводних.
Цікаво відзначити, що клітини зародків ссавців (розд. 6.6.1), незважаючи на те що у названих тварин яйцеклітини мають малу кількість жовтка, а дроблення повне, в фазі гаструляції
зберігають переміщення, властиві зародкам плазунів і птахів. Це підтверджує уявлення про походження ссавців від предковой групи, у якій яйця були багаті жовтком.
Гаструляция характеризується різноманітними клітинними процесами. Триває митотичне розмноження клітин, причому воно має різну інтенсивність в різних частинах зародка. Разом з тим найбільш характерна риса гаструляціі полягає в переміщенні клітинних мас. Це призводить до зміни будови зародка і перетворення його з бластули в гаструлу. Відбувається сортування клітин по їх приналежності до різних зародковим листками, всередині яких вони «дізнаються» один одного.
На фазу гаструляції припадає початок цітодіфференціровкі, що означає перехід до активного використання біологічної інформації власного геному. Одним з регуляторів генетичної активності є різний хімічний склад цитоплазми клітин зародка, що встановився внаслідок овоплазматічної сегрегації. Так, ектодермальние клітини земноводних мають темний колір через пігменту, який потрапив в них з анімального полюса яйцеклітини, а клітини ентодерми - світлий, так як відбуваються з вегетативного полюса яйця.
Під час гаструляції дуже велика роль ембріональної індукції. Показано, що поява первинної смужки у птахів - результат індукційного взаємодії між гіпобласт і Епібласт. Гіпобласт властива полярність. Зміна положення гіпобласта по відношенню до Епібласт викликає зміна орієнтації первинної смужки.
Детально про всі перераховані процеси розказано в розд. 8.2. Слід зауважити, що такі прояви цілісності зародка як детермінація, ембріональна регуляція і інтегрованість притаманні йому під час гаструляції в тій же мірі, як і під час дроблення (див. розд. 8.3).