| Головна |
| «« | ЗМІСТ | »» |
|---|
Зміни спадкової програми статевих клітин людини призводять до народження потомства з різними спадково зумовленими хворобами, в залежності від рангу мутацій - генними або хромосомними.
Різні генні мутації по-різному позначаються на життєздатності організму, причому в разі їх рецессивности вони можуть довго не проявлятися фенотипно у нащадків. Хромосомні перебудови і геномні мутації призводять до виражених відхилень у розвитку і часто є причиною загибелі організму на різних стадіях його онтогенезу, зазвичай в ранньому ембріогенезі. Значною мірою саме цими мутаціями визначається високий відсоток (15%) переривання діагностованих вагітностей.
Триплоїд плода, як правило, призводять до переривання вагітності на ранніх стадіях, проте описано дуже невелике число випадків живорождения Триплоїд. анеуплоідія за різними хромосомами зустрічається як в матеріалі абортусов, так і у народжених дітей. Деякі анеуплоідіі несумісні з життям. Так, трісо- мія по 16-й хромосомі виявляється тільки в матеріалі абортусов. У той же час у людини відомі синдроми, пов'язані з аномаліями числа хромосом, що характеризуються різним ступенем життєздатності.
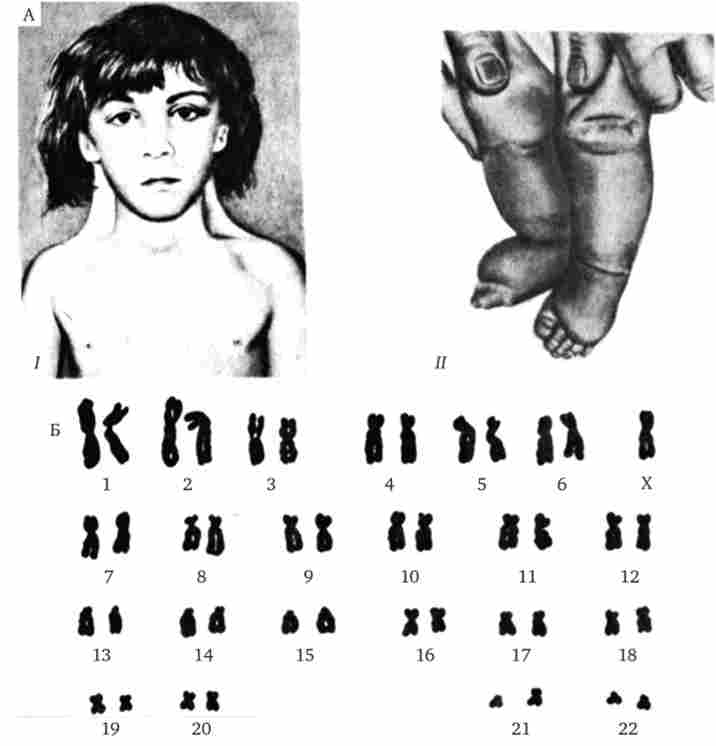
Найбільш частим хромосомним захворюванням у людини є синдром Дауна, обумовлений трисомія по 21-й хромосомі, що зустрічається з частотою 1-2 на 1000 (рис. 4.3). Приблизно в 60%
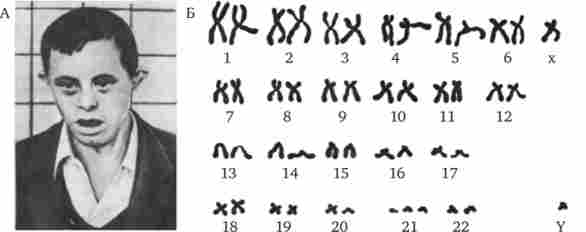
Мал. 4.3. Синдром трисомії 21 (синдром Дауна).
А - зовнішній вигляд хворого; Б - каріотип хворого
випадків трисомія 21 є причиною загибелі плоду, близько 30% народжених вмирає на першому році життя. Ще 46% не переймається 3-річний рубіж, проте іноді люди з синдромом Дауна доживають до значного віку (рис. 4.4), хоча в цілому тривалість їхнього життя скорочена. Застосування ефективних протимікробних препаратів дозволяє дещо збільшити тривалість життя таких хворих. Трисомія 21 може бути результатом випадкового нерасхожде- ня гомологічниххромосом в мейозі. Поряд з цим відомі випадки регулярної трисомії, пов'язаної з транслокацией 21-ї хромосоми на іншу - 21, 22, 13, 14 або 15-ю хромосому (рис. 4.5).

Мал. 4.4. Жінка з синдромом Дауна у віці 38 років
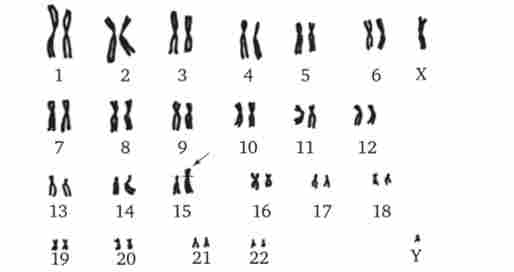
Мал. 4.5. Каріотип при Транслокаційний синдромі Дауна (одна 21-я хромосома приєднана до 15-й хромосомі - зазначено стрілкою)
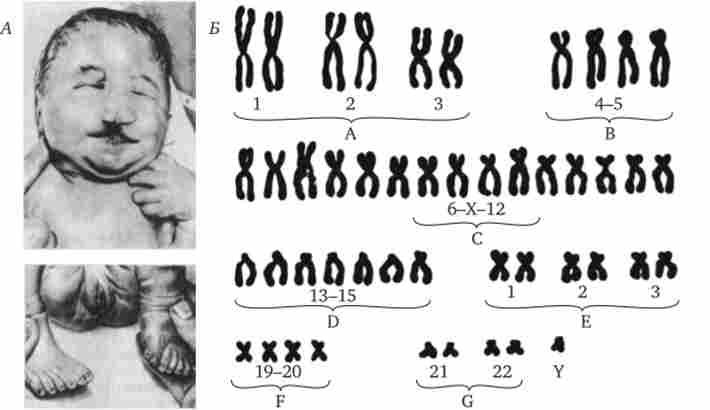
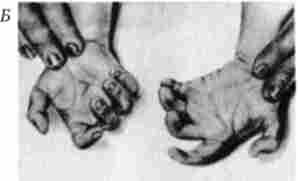
Мал. 4.6. Синдром трисомії 13 (синдром Патау).
А - зовнішній вид хворого; Б - каріотип хворого з трисомія в групі D; / - аномалії особи, II - двостороння полісіндактілія стоп
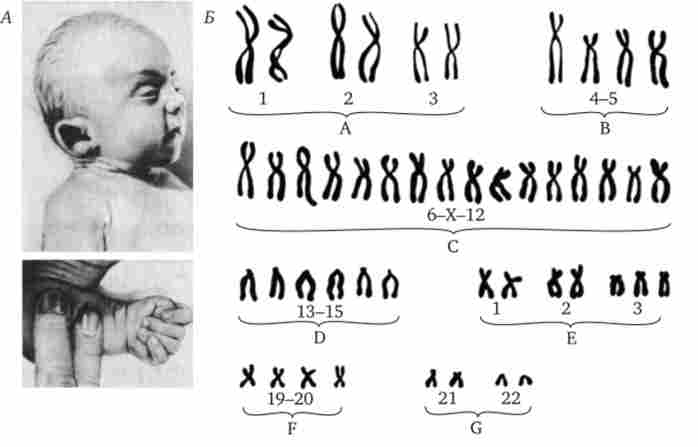
Мал. 4.7. Синдром трисомії 18 (синдром Едвардса).
А - зовнішній вид хворого; Б - каріотип хворого при трисомії в групі Е;
I - черепно-лицьові аномалії, II- характерне розташування пальців на кистях хворого
Серед інших аутосомних трисомій відомі трисомії по 13-й хромосомі - синдром Патау (Рис. 4.6), а також по 18-й хромосомі - синдром Едвардса (Рис. 4.7), при яких життєздатність новонароджених різко знижена. Вони гинуть в перші місяці життя через множинних вад розвитку. Застосування методів диференціального фарбування хромосом дозволило відкрити три нових синдрому, зумовлених трисомиями по 8, 9 і 22-й хромосомах, при яких також спостерігаються важкі комплексні вади розвитку (рис. 4.8).

Мал. 4.8. Синдром трисомії 8.
А - зовнішній вид хворого;
Б - контрактури в межфаланго- вих суглобах кистей
Досить часто у людини зустрічаються анеуплоідіі за статевими хромосомами (Рис. 4.9-4.11). На відміну від анеуплоідіі по ауто- сомам дефекти розумового розвитку у хворих виражені не так чітко, у багатьох воно в межах норми, а іноді навіть вище середнього. Разом з тим у них постійно спостерігаються порушення розвитку статевих органів і гормонозалежного зростання тіла. Рідше зустрічаються пороки розвитку інших систем. Щодо сприятливі наслідки збільшення числа Х-хромосом, мабуть, пов'язані з можливістю компенсації дози відповідних генів завдяки природній генетичної інактивації цих хромосом, а також мозаїчному характеру такої інактивації.
Мал. 4.9. Синдром моносомии X (ХО-синдром, синдром Шерешерского - Тернера).
А - зовнішній вигляд хворої; Б - каріотип жінки з синдромом ХО; / - виражена трапецієвидна шийна складка, широка грудна клітка, широко розставлені, слаборозвинені соски молочних залоз, II - характерні лімфатичні
набряки на ногах
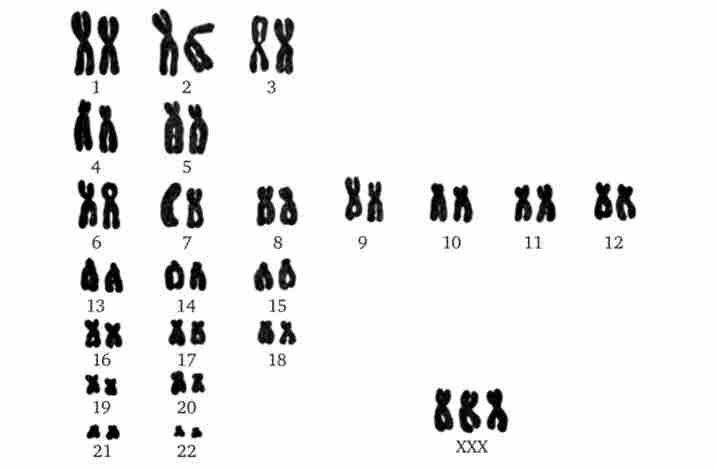
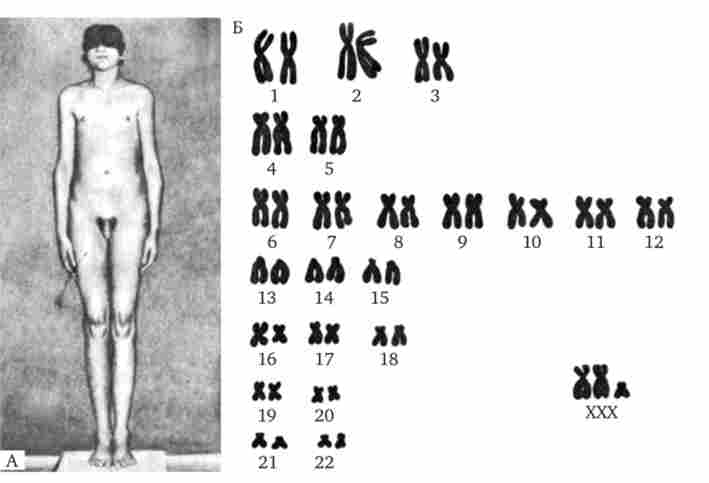
Мал. 4.10. Каріотип жінки з синдромом трисомії X Мал. 4.11. Синдром Клайнфельтера.
А - зовнішній вид хворого (характерний високий зріст, непропорційно довгі кінцівки); Б - каріотип хворого (XXY)
Серед анеуплоідних синдромів за статевими хромосомами моно- сомія X (ХО) (Синдром Шерешевського - Тернера) Зустрічається багато рідше, ніж трисомия X, синдром Клайнфельтера (XXY, XXXY), а також
XYY, що вказує на наявність сильного відбору проти гамет, що не містять статевих хромосом, або проти зигот ХО. Це припущення підтверджується досить часто спостерігається моносомією X серед спонтанно абортованих ембріонів. У зв'язку з цим допускається, що вижили зиготи ХО є результатом не мейоті- чеського, а митотичного нерасхожденія, або втрати Х-хромосоми на ранніх стадіях розвитку (див. Рис. 4.9). Моносомії Y0 у людини не виявлено.
Організми з анеуплоідіі за статевими хромосомами при наявності Y-хромосоми розвиваються за чоловічим типом і фенотипично дають синдром Клайнфельтера (див. Рис. 4.11). Це є ще одним свідченням на користь розташування фактора, визначає чоловічу тип розвитку в Y-хромосомі.
З синдромів, пов'язаних зі структурними аномаліями хромосом, відомий транслокаційний синдром Дауна (Див. Рис. 4.5), при якому число хромосом в каріотипі формально не змінено і дорівнює 46, так як додаткова 21-а хромосома транслоцироваться на одну з акроцентрічних хромосом. При транслокації довгого плеча 22-ї хромосоми на 9-ю розвивається хронічний мієлолейкоз. При делеции короткого плеча 5-ї хромосоми розвивається синдром котячого крику, при якому спостерігаються загальне відставання в розвитку, низька маса при народженні, місяцеподібне обличчя з широко розставленими очима і характерний плач дитини, що нагадує котяче нявкання, причиною якого є недорозвинення гортані (рис. 4.12).
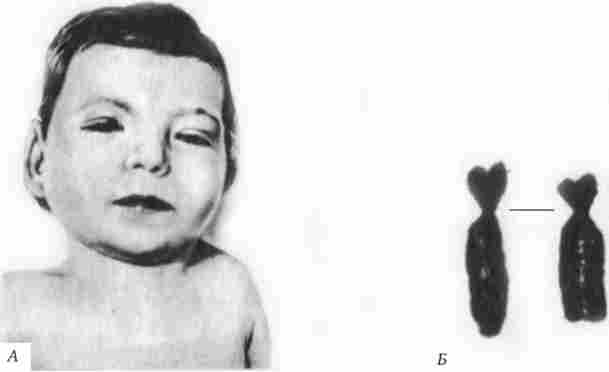
Мал. 4.12. Синдром хромосоми 5р (синдром «котячого крику»).
А - зовнішній вид хворого; Б - ділення короткого плеча 5-ї хромосом
У носіїв деяких періцентрічних інверсій нерідко спостерігаються аномалії у вигляді розумової відсталості тій чи іншій мірі і пороків розвитку. Досить часто такі перебудови спостерігаються в 9-й хромосомі людини, однак вони суттєво не впливають на розвиток організму.
Таким чином, порушення спадкової програми організму, що розвивається з аномальних гамет, або мозаїцизм його клітин, пов'язаний з соматичними мутаціями, є причиною або загибелі організму, або більш-менш вираженого зниження його життєздатності.