| Головна |
| «« | ЗМІСТ |
|---|
Мутації різного рангу (генні, хромосомні або геномні), що виникають в соматичних клітинах організму, успадковуються нащадками цих клітин і роблять організм мозаїка, т. е. особиною зі змішаними популяціями клітин. У розд. 3.6.5.1 і 3.6.5.2 розглянуті приклади природного мозаїцизму жіночого організму по активно функціонуючим в його клітинах Х-хромосомами і пов'язане з цим явище аллельного виключення, коли в різних клітинах організму експресуються різні аллели Х-зчеплених генів.
Наприклад, у жінки - гетерозиготной носії рецесивного алеля гемофілії - ступінь порушення згортання крові залежить від співвідношення відповідних клітин з генетично інактивованих Х-хромосомами, що несуть нормальний або мутантний аллель.
Нерідко у людини зустрічається мозаїцизм по геномних мутацій, пов'язаний з порушенням розбіжності хромосом при мітозі. Наприклад, в разі синдрому Дауна (трисомія по 21-й хромосомі) мозаицизм зустрічається з частотою 2 на 48 пацієнтів, а в популяції їх частота дорівнює 1 на 31 000. Чим раніше в ході розвитку організму відбувається порушення поділу соматичних клітин, що супроводжується нерозходженням дочірніх хромосом до полюсів ахроматіно- вого веретена, тим більш вираженою буде симптоматика захворювання, що викликається даної анеуплоідіі. Порушення мітозу на більш пізніх стадіях індивідуального розвитку призводить до локального мозаїцизму, який може не супроводжуватися вираженими відхиленнями від норми. У цьому випадку найбільш небезпечним є мозаицизм клітин генеративних тканин, з яких з досить великою ймовірністю організм може утворювати гамети з аномальним числом хромосом.
Іноді виникають соматичні мутації є причиною появи злоякісних новоутворень. На рис. 4.2 представлена послідовність подій, що призводять до утворення таких пухлин. Як правило, в основі лежить пошкодження ДНК, що викликається внутрішніми факторами (порушенням процесів реплікації, репарації або рекомбінації) або зовнішніми впливами (іонізуючоїрадіацією, хімічними мутагенами або вірусами). Одним з результатів такого пошкодження ДНК може виявитися поява клону клітин, що володіє дефектами регуляції клітинного розмноження, що призводить до пухлинного росту.
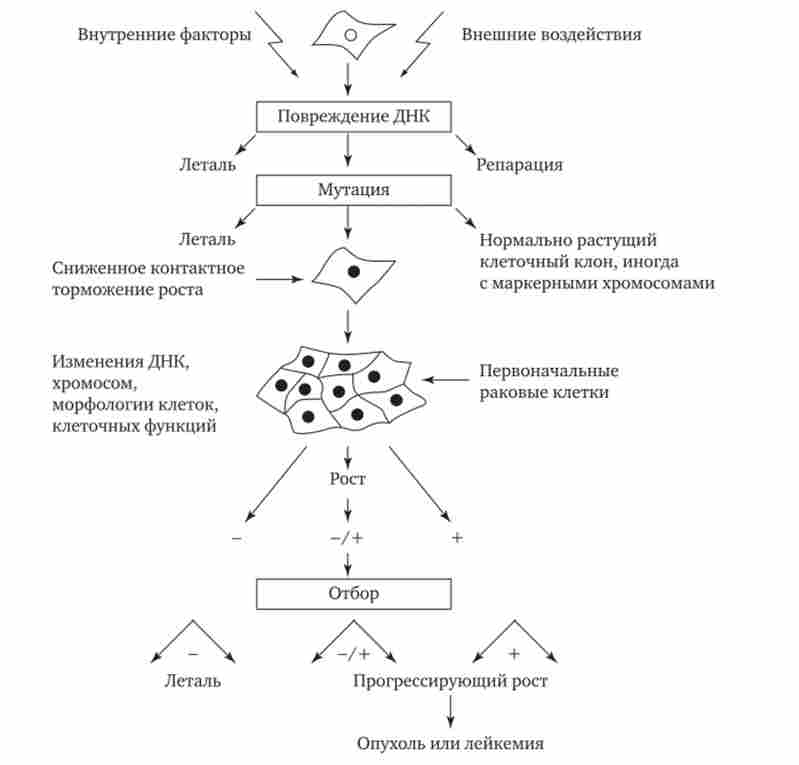
Мал. 4.2. Соматичні мутації як причина злоякісного переродження тканин
Причиною злоякісного розростання тканини можуть бути також порушення мітозу і нерівноцінне розподіл хромосом між дочірніми клітинами з виникненням анеуплоідіі або хромосомних аберацій. Це викликає або загибель клітин, або призводить до появи клонів, здатних до неконтрольованого зростання. У злоякісні утворення зазвичай зустрічаються субклони, що мають різні каріотипи, що свідчить про множинних аномаліях мітозу в клітинах пухлин.
Так як в основі злоякісного переродження тканин може лежати зміна спадкового матеріалу клітин, стає очевидною важлива роль мутагенних чинників в процесі виникнення пухлин. Одним з таких мутагенних чинників є віруси, які можуть індукувати в хромосомах ту чи іншу мутаційна зміна. Серед пухлин людини вірусне походження має лімфома Беркітта.
Розумінню молекулярно-біологічних механізмів пухлинного росту сприяло відкриття так званих онкогенов, викликають злоякісне переродження клітин і входять до складу геному ретровірусів. Геном ретровируса складається з одноцепочечной РНК і містить ген зворотної транскриптази. При проникненні вірусу в клітину-господаря під контролем ферменту зворотної транскриптази утворюються множинні копії генетичної інформації вірусу, але вже у вигляді дволанцюгової ДНК, які вбудовуються в ДНК клітини-господаря. Функціонування таких ДНК-копій у складі генома клітини призводить до синтезу вірусних РНК і білків, причому вірусний онкоген (v-onc) трансформує клітку-господаря в пухлинну.
Використання ДНК-зондів на основі ретровірусних онкогенів виявило їх гомологію деяким власним нуклеотидних послідовностей генома клітин. Ці ділянки отримали назву протООНкогенов, або клітинних онкогенів (З-ОПС). ПротООНкогени беруть участь в контролі клітинного росту, але в звичайному стані не призводять до пухлинної трансформації. Їх мутантні аллели, стимулюючи мітоз, можуть викликати ріст пухлин. Іноді активація клітинного онкогена обумовлюється одиничної точковой мутацією в ньому. В інших випадках немає необхідності в такій мутації, так як трансформуючий ефект спостерігається при приєднанні онкогена до фрагментів ДНК, які мають сильними промоторних властивостями. У таких ситуаціях слід допустити, що протООНкогенах притаманні властивості транспозони, або «стрибаючих генів».
Припускають, що вірусні онкогени на якомусь етапі еволюції походять від клітинних онкогенів, інтегрованих в геном вірусу. Причини, що зумовлюють велику активність, що трансформує вірусних онкогенів в порівнянні з клітинними, до теперішнього часу не цілком ясні.
У пухлинних клітинах часто спостерігаються хромосомні аномалії, причому деякі пухлини відрізняються наявністю специфічних хромосомних дефектів.
Встановлено, що онкогени нерідко виявляються в безпосередній близькості від точок розривів, що відбуваються при опухолеспеціфічних хромосомних перебудовах. Це підтверджує їх роль в злоякісної трансформації клітин.