| Головна |
| «« | ЗМІСТ | »» |
|---|
Розрізнення висоти звуку пов'язане з особливостями коливань основний мембрани в різних частинах равлики. Ця різниця пояснюється тим, що основна мембрана в міру віддалення від овального вікна і наближення до гелікотреми ширшає і її пластичність зростає. Завдяки цьому мембрана діє як аналізатор частоти звуку: від частоти звуку залежить, в якій частині основної мембрани будуть виникати коливання (рис. 5.23). Ділянки первинної слухової кори сприймають звукові сигнали певної частоти, причому їх організація повторює організацію рецепторного апарату равлики. Тривалість звукового стимулу кодується тривалістю активності нейронів, а інтенсивність - частотою імпульсації.
При використанні різних тонів було показано, що по-лосковие клітини в різних частинах равлики краще налаштовані на якусь певну частоту, що може бути пов'язано із спеціальними властивостями покривної мембрани. Волосся-кові клітини мають властивість двухтоновий придушення. Воно полягає в наступному: якщо при реакції клітин на тон характеристичної частоти додати інший тон, частота якого вище або нижче, то реактивність клітин в області характеристичної частоти знижується. Це явище - посилення контрасту - одне із загальних властивостей сенсорних си
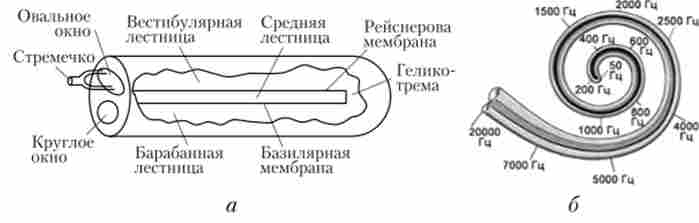
Мал. 5.23. Схема равлики в розгорнутому вигляді (а); відповідні зміни частоти коливань основний мембрани уздовж довжини равлики (Б)
стем, що існують на всіх їх рівнях і пояснюється синаптичними механізмами, наприклад латеральним гальмуванням. Посилення контрасту проявляється вже на рівні сенсорної периферії. В процесі перетворення звуку беруть участь два фільтри: перший фільтр - хвиля, що біжить, другий - «загострює» механізм, пов'язаний з рухом основний мембрани і зміщенням волосків.
У слуховому нерві людини всього 25 000 волокон. З них 90% волокон пов'язано з внутрішніми волосовими клітинами, які складають лише 20% всіх рецепторних клітин. Інша частина нервових волокон пов'язана з великим числом зовнішніх волоскових клітин. Таким чином, і на периферії, і в центральних відділах ми зустрічаємося з множинною іннервацією невеликого числа клітин (конвергенцією), Що забезпечує надійність передачі сенсорної інформації (див. Рис. 5.2). дівергентние відносини, характерні для зовнішніх волоскових клітин, навпаки, забезпечують активність декількох сусідніх клітин при активації однієї зовнішньої волоскової клітини.
Вважається, що основний потік імпульсів в слуховому нерві пов'язаний з внутрішніми волосовими клітинами, з якими контактує 90% аферентних волокон; зовнішні волоскові клітини, тісно контактують з текторіальная мембраною, підсилюють звуковий сигнал і тим самим збільшують можливість функціональних контрастів. Синаптичні контакти і сенсорні волокна забезпечують точну передачу характеристичних кривих рецепторних клітин. Важливим є той факт, що нервові імпульси виникають в нервовому волокні синхронно при частоті 1000 коливань в секунду (1 кГц). Проводити велику частоту імпульсів волокна не здатні. Як і в інших сенсорних системах, частота імпульсів в аферентних волокнах кодується і положенням рецепторних утворень на основній мембрані. Частотою імпульсів в нерві кодується інтенсивність стимулу. Незважаючи на малу кількість сенсорних виходів, обидва види пристосувань забезпечують гострий слух.