| Головна |
| «« | ЗМІСТ | »» |
|---|
Найбільш добре вивченим хімічним синапсом є нервово м'язову з'єднання, або кінцева платівка. Однак механізми, розглянуті на цьому прикладі, застосовні і до всіх інших хімічних синапсах в нервовій системі.
Почнемо розгляд механізму передачі інформації з надходження ПД до синаптичних закінчення (рис. 5.3). Оскільки синаптична щілину в хімічному синапсі має досить великі розміри, для ефективної передачі імпульсу до наступної клітці необхідний механізм посилення. Це наявність хімічної речовини - медіатора (посередника). У випадку з кінцевою пластинкою цим медіатором є речовина ацетилхолін.
Експериментально було показано, що медіатор в синапсі міститься в спеціальних бульбашках - везикулах, а не просто розчинений в цитоплазмі. Бульбашки формуються шляхом впячивания мембрани пресинаптичного закінчення. Бувають бульбашки облямовані і гладкі. Ацетилхолін синтезується в тілі клітини, а в самому закінченні. Там знаходиться ферментативна система синтезу цієї речовини.
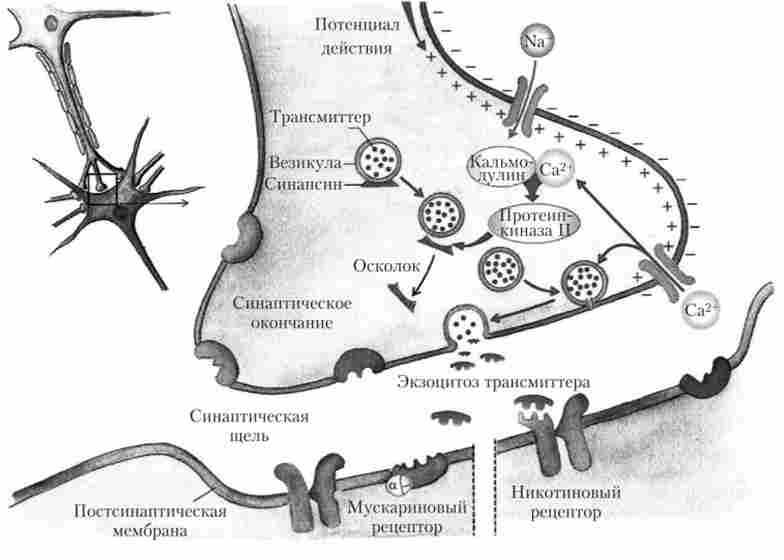
Мал. 5.3. Процеси передачі сигналу в хімічному синапсі1
Ацетилхолін широко поширений не тільки в організмі людини, але міститься також в жалких капсулах кропиви і жалких клітинах кишковопорожнинних тварин.
При дослідженні діяльності нервово-м'язового синапсу проводили такі експерименти. Вводили мікроелектрод в м'язову клітку в області синапсу (під постсинаптичної мембраною) і реєстрували зміна її потенціалу. Виявилося, що в стані спокою, т. Е. Без нервового імпульсу, ПП м'язової клітини не виражається прямою лінією. Періодично спостерігаються сплески рівня мембранного потенціалу (невелика за амплітудою деполяризация) з частотою приблизно один раз в секунду. Ці потенціали були названі мініатюрні потенціали кінцевої пластинки (МПКП). Вивчення роботи синапсу за допомогою електронного мікроскопа дозволило зв'язати кожен такий МПКП з виділенням порції (кванта) медіатора, яка міститься в одному бульбашці. так виникла квантова теорія вивільнення медіатора. Вона постулює, що в кожному хімічному синапсі медіатор вивільняється квантами, або порціями, які дорівнюють змістом медіатора в одному бульбашці.
Таким чином, навіть в стані спокою періодично відбувається виділення одного кванта ацетилхоліну в синаптичну щілину, що призводить до виникнення в м'язовій клітці зсуву мембранного потенціалу в позитивну сторону, т. Е. До появи МПКП. У стані возбуж-
дення відразу збільшується ймовірність виділення медіатора (одночасно вивільняється більше квантів) і в м'язовій клітці реєструється вже не мініатюрний, а просто потенціал кінцевої пластинки (1IKII). Коли виділяється відразу велику кількість квантів медіатора, то МПКП від кожного окремо складаються, і сума МПКП утворює ПКП. Якщо медіатора виділилося досить багато, то амплітуда Г1КГ1 досягне порогового значення і розвинеться ПД м'язової клітини. Результатом ПД м'язової клітини є скорочення м'язових волокон.
Розглянемо процеси, що відбуваються в синапсі, які призводять до появи ПКП і в результаті - до м'язового скорочення. Отже, по нервовому волокну до закінчення приходить ПД (збудження) і починається деполяризація пресинаптичної мембрани. У цей час змінюється заряд на мембрані (внутрішня сторона змінює знак з негативного на позитивний) і запускається вивільнення іонів Са++ з внутрішньоклітинних депо (ендоплазматичнийретикулум). Ці процеси призводять до того, що бульбашки з медіатором починають у великій кількості підходити до пресинаптичної мембрани і вбудовуватися в неї (процес екзо цитоза), в результаті чого ацетилхолін виходить в синаптичну щілину. Далі ацетилхолін рухається до постсинаптичні мембрані, т. Е. Вже до мембрани м'язової клітини. На постсинаптичні мембрані є спеціальні білкові освіти - рецептори, з якими ацетилхолін і зв'язується. Холінорецептори представляють собою структури, пов'язані з іонними каналами, тому при зв'язуванні медіатора зі своїм рецептором відбувається відкривання каналу для будь-якого іона. У випадку з ацетилхоліном - це канали для натрію і калію.
Таким чином, при взаємодії ацетилхоліну зі своїм рецептором відбувається відкривання іонного каналу в мембрані м'язової клітини, виникають потік іонів натрію всередину клітини і деполяризація мембрани м'язової клітини. Якщо струм і, отже, деполяризация досить сильні, то в м'язовій клітці виникне ПД.
Для нормальної роботи синапсу, зокрема для забезпечення передачі параметрів (частоти і патерну) приходить до синапси інформації, необхідно, щоб медіатор досить швидко віддалявся з постсинаптичних рецепторів, т. Е. З синаптичної щілини. Це забезпечується, крім незначного витоку медіатора з області синаптичної щілини, перш за все, двома механізмами. По-перше, це ферментативна інактивація медіатора шляхом розщеплення його активної молекули на неактивні метаболіти (продукти розпаду). У нервово-м'язовому синапсі цю функцію в синаптичної щілини виконує фермент ацетилхолінестерази, яка розщеплює ацетилхолін на складові елементи - холін і оцтову кислоту (ацетат). Другим способом усунення медіатора з щілини є зворотне захоплення медіатора, коли його молекули надходять в пре- синаптічеськоє закінчення. Так відбувається, наприклад, в норадренергіче- ських синапсах як в ЦНС, так і на периферії (в складі симпатичної нервової системи). Це забезпечує не тільки усунення активної речовини з щілини, але і більш економне витрачання медіатора в синапсі.
У разі кінцевої пластинки зворотному захопленню піддається не сам аце- тілхоліна, а що утворюється при його розпаді холін, який в подальшому знову використовується в синаптичному закінчення для синтезу ацетилхоліну. Для синтезу нового ацетилхоліну в закінченні аксона існує інший фермент - ацетілхолінтрансфераза.