| Головна |
| «« | ЗМІСТ | »» |
|---|
Гормони здатні надавати ряд фізіологічних процесів.
Метаболічну, пов'язане зі зміною обміну речовин. Більшість гормонів беруть участь в регуляції обміну речовин шляхом зміни активності ферментативних систем в тканинах. Для деяких гормонів вплив на обмінні процеси є основною функцією. Наприклад, інсулін, глюка- гон і адреналін безпосередньо регулюють вуглеводний обмін; глюкокортикоїди стимулюють утворення вуглеводів із продуктів розпаду білків, мінералкортікоіди впливають на вміст натрію і калію в організмі, а гормон околощітовідной залози регулює обмін кальцію і фосфору. Соматотропний гормон гіпофіза стимулює синтез білка і витрачання вуглеводів і жирів. Вплив на рівень енергетичних процесів гормонів щитовидної залози реалізується за рахунок посилення розщеплення білків, вуглеводів і жирів. Вплив статевих гормонів проявляється в посиленні синтезу білків (особливо м'язових) і інтенсивному витрачання жиру і зміні мінерального обміну.
Морфогенетичне, пов'язане зі зміною диференціації клітин і тканин, зростанням і метаморфозом. Добре відомо дію гормонів щитовидної залози на метаморфоз пуголовків; порушення її функцій призводить до відчутних порушень росту і розвитку молодняку більшості тварин. Гонадотропні гормони гіпофіза стимулюють зростання, розвиток і диф- ференціровку клітин статевих залоз. Початок продукції гормонів статевими залозами обумовлює розвиток статевих органів і поява вторинних статевих ознак.
Кінетичне, або пусковий, вплив, що викликає діяльність ефекторних структур. Ендокрінокінетічне дію характерно для тропних гормонів гіпофіза, які необхідні для «включення» вироблення гормонів щитовидної залози, статевих залоз і гормонів кори надниркової залози. Крім того, «пусковий» вплив деяких нейрогіпофізарного гормонів полягає в ініціації скорочень гладкої мускулатури матки, міоепітелія молочних залоз, гладкої мускулатури судин.
Корегуючий, пов'язане зі зміною рівня інтенсивності функцій організму або його органів, що працюють і без участі гормонального впливу. При цьому відбувається посилення або ослаблення фізіологічних процесів. Так, адреналін прискорює ритм і збільшує силу серцевих скорочень, але пригнічує скоротливу активність мускулатури шлунково-кишкового тракту.
Ключовий етап в реалізації фізіологічної дії гормону на клітину-мішень - це його зв'язування зі специфічним білком-рецептором, який служить розпізнає посередником гормонального ефекту. Наявність рецептора - необхідна умова розвитку ендокринної функції, і, якщо в клітці немає рецепторів, гормон не здатний впливати на неї.
У загальному вигляді рецептор для будь-якого з гормонів складається з трьох просторово відокремлених структур:
. здійснює виборчий прийом гормонального сигналу за рахунок специфічного і оборотного зв'язування гормону;
здійснює перетворення зовнішнього гормонального сигналу у внутрішньоклітинний сигнал;
відповідальної за ініціацію регуляторних ефектів гормону за рахунок взаємодії гормонорецепторного комплексу з різними акцепторними ділянками клітини (рис. 12.1).
Таким чином, рецептор - це така хімічна структура відповідної тканини-мішені, яка має високоспецифічні ділянки для зв'язування гормональних сполук, причому в результаті цього зв'язування ініціюються наступні біохімічні реакції, необхідні для здійснення кінцевого ефекту даного гормону.
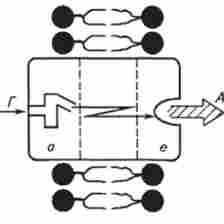
Виходячи з локалізації рецепторів, характеру акцепторних ділянок і особливостей гормонзавісімих ефектів, рецепцію поділяють на внутрішньоклітинну і мембранну. внутрішньоклітинна рецепція характерна для стероїдних і тиреоїдних гормонів. Гормон вільно проникає через плазматичну мембрану всередину клітини і взаємодіє з цитозольними, ядерними та іншими рецепторами. мембранозв'язані рецептори пов'язують гормони на поверхні клітин, і гормональні ефекти розвиваються завдяки освіті внутрішньоклітинних посередників (медіаторів) в результаті взаємодії гормонорецепторного комплексів з мембран-
Мал. 12.1. Модель функціонально-структурної організації рецепторной молекули:
а - гормоносвязиваюшій ділянку; е - еффекторний ділянку, звивиста лінія - ділянка сполучення а і е А - акцептор, Г - гормон ними акцепторними структурами. Як правило, мембранним акцептором служить або аденилатциклаза, що змінює рівень внутрішньоклітинного циклічного аденозинмонофосфату (цАМФ), або неелектрогенние кальцієві канали, що змінюють внутрішньоклітинну концентрацію іонізованого кальцію, які специфічні протеази, здатні змінити внутрішньоклітинний вміст специфічних глікопептидів.
Існує і можливість внутрішньоклітинного дії пов'язаного з мембраною гормону. Мембранні рецептори зазвичай асиметрично вбудовані в плазматичну мембрану, причому зв'язує гормонузнающій фрагмент їх молекул жорстко орієнтований до зовнішньої поверхні і звернений до позаклітинного простору. Взаємодіє з акцептором частина рецепторной молекули (або рецепторного комплексу) орієнтована в протилежному напрямку і звернена до цитоплазми клітини. Ця жорстка орієнтація рецепторів забезпечує століття-торность трансмембранної передачі гормональної інформації всередину клітин. Локалізуючись трансмембранно, гідрофобна частина мембранних (поверхневих) рецепторів виявляється зануреною в біслойную липидную мембрану, в той час як зв'язують і виконавчі ділянки знаходяться у водній фазі. Незважаючи на відносну жорсткість фіксації таких рецепторів в мембрані, вони здатні переміщатися, здійснюючи поздовжні і поперечні руху, полегшуючи тим самим «пошук» рецептором гормону. Можлива рухливість рецепторних молекул дозволяє здійснити клітці такі процеси, як агрегування і интернализация рецепторів, що зв'язали гормональні молекули. В основі цього процесу лежить спеціальний процес - адсорбтівний ендоцитоз. Дифузно розподілені по клітинної поверхні рецептори після зв'язування гормону набувають здатність до рухів в латеральної площині. Зближуючись, вони об'єднуються (кластеризуються) в особливих ділянках плазматичної мембрани - облямованих ямках - місцях почала інтерналізації. За даними електронної мікроскопії, облямовані ямки є поглиблення в клітинній мембрані з боку цитоплазми, вистелені шаром щетінкоподобного білка - клатріна, що виконує функцію своєрідного каркаса (клатріновая кошик). Завантажені кластерами гормонорецепторного комплексів облямовані ямки поступово заглиблюються, замикаються і відриваються від мембрани, перетворюючись в вільно плаваючі по цитоплазматичних простору везикули. В ході переміщення по цитоплазмі відбувається втрата клатрінового покриття, гладкі везикули об'єднуються, формуючи рецептосоми, які здатні зливатися з лізосомами або структурами апарату Гольджі. Освіта комплексної ендолізосомальной везикули призводить до ферментативному поділу комплексу гормон-рецептор,
Ріс.12.2. Схема рецепторопосрелован- ного енлоцітоза:
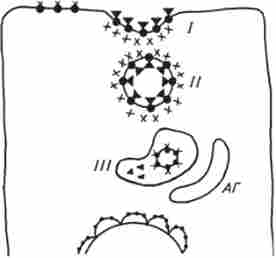
/ - облямована ямка; // - ендосомамі; /// - мульти везикулярне ендосоми; АГ апарат Гольджі
причому «проник» в складі ендосоми гормон здатний зв'язуватися з внутрішньоклітинними структурами, а рецептор має можливість знову вбудуватися в плазматичну мембрану (ре- циклировать) (рис. 12.2).