| Головна |
| «« | ЗМІСТ | »» |
|---|
Принципові риси будови клітин тварин, рослин, грибів однакові. Їх спільна риса - компартменталізація. Цей термін означає підрозділ клітини на ядро, що містить хроматин і одне або кілька ядерець, і цитоплазму, в якій розрізняють мітохондрії, пластиди (у рослин) і деякі інші самовідтворюються орга Неллі клітини. Крім того, в цитоплазмі еукаріот є органели, які постійно присутні в ній, але не мають здатність до самовідтворення. Це апарат Гольджі, вакуолі, лізосоми.
Клітини бактерій (прокаріотів) побудовані по іншому типу: вони не мають морфологічно оформлених самовідтворюються органел, за винятком нуклеоида - аналога ядра клітини еукаріот.
Будь-яка активно ділиться клітина зазнає ряд послідовних змін, з яких складається клітинний цикл. За оптимальних умов тривалість клітинного циклу у різних організмів неоднакова.
Клітинний цикл складається з чотирьох періодів: Пресинтетичний (Gi), періоду синтезу ДНК (5), постсинтетичному (Gy і мітозу (Л /). Власне мітоз захоплює не більше 1 / 7-1 / 10 всього клітинного циклу, складаючи лише невелику частину його .
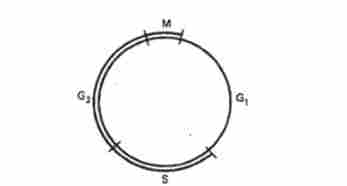
Мал. 3.1. Схема клітинного циклу
У яка покоїться клітці на стадії інтерфази розрізняють сферичне ядро, оточене двошаровою ядерною мембраною з порами діаметром близько 40 мкм. Хромосоми в цій стадії знаходяться в ядрі в розправленому, розтягнутому стані і практично невидимі в світловий мікроскоп до початку мітозу Проте, вже на перших етапах розвитку цитології були отримані факти, що вказують на безперервність існування хромосом в часі. Про це свідчить сталість форми (збереження індивідуальності) хромосом. Ще в 1888 р Т. Бовери показав, що в перших мітозів заплідненого яйця аскариди хромосоми в профазі з'являються в тому ж місці, де вони опинилися в телофазе попереднього мітозу. У наші дні це було підтверджено подружжям Байєр (1957- 1958 рр.), Які застосували мікрокінозйомок для спостереження митозов в ендоспермі лілійних.
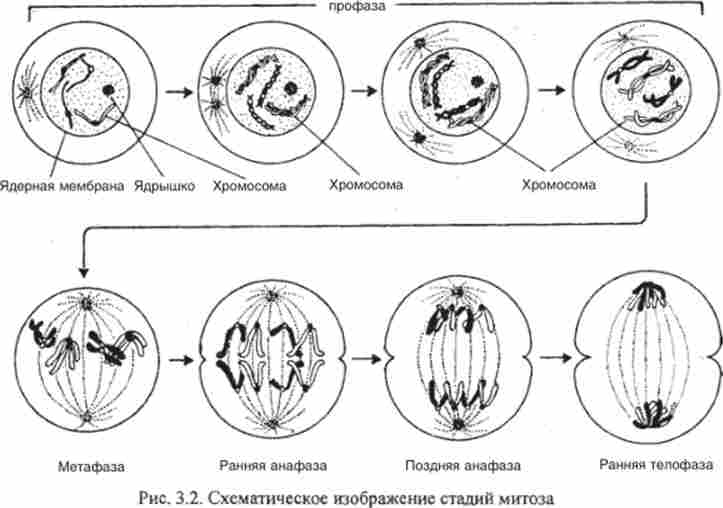
Профаза. Хромосоми спірапізуются, скорочуючи і потовщені. Завдяки цьому вони стають помітними в світловий мікроскоп. Уже в ранній профазі видно, що кожна хромосома складається з двох хроматид. Це результат реплікації, що сталася в інтерфазі - на стадії S клітинного циклу. Хроматиди тісно асоційовані по довжині і перекручені. До кінця профази ця асоціація слабшає. Зникають ядерця, розчиняється ядерна мембрана, і хромосоми опиняються в цитоплазмі. Цей момент позначає завершення профази.
На початку профази хромосоми рівномірно розподіляються по всьому ядру, а до кінця профази наближаються до ядерної мембрані. Перед розчиненням ядерної мембрани хромосоми майже повністю спіралізують. За період профази вони скорочуються до 1/25 довжини в порівнянні з ранньої Профазі.
Прометафаза. Після розчинення ядерної мембрани хромосоми рухаються у напрямку до екватора. Прометафаза завершується, коли хромосоми досягають екваторіальній площині.
Мстафаза. З'являються неокрашіваемие нитки ахроматинового веретена. У тваринних клітинах видно зіркоподібні фігури біля полюсів. Вони утворюються навколо центріолей, що розійшлися до полюсів клітини. Хромосоми (кожна з двох хроматид) шикуються в площині екватора, утворюючи так звану метафазну пластинку. Довгі хромосоми як правило зігнуті F-образно. При цьому вістря V вказує на центр метафазної пластинки, і на ньому видно неокрашенная перетяжка - центромера, до якої прикріпляється нитка ахроматинового веретена і таким чином забезпечує рух хромосоми. Центромери шикуються в екваторіальній площині, а плечі досить довгих хромосом як би звисають до того чи іншого полюсу.
Метафазної платівка - своєрідний паспорт організму. Число і форма хромосом, які спостерігаються в метафазі, характеризують каріотип виду. Чітко виявляється парність хромосом. Парні, або гомологічні, хромосоми зазвичай абсолютно однакові по довжині, розташуванню центромери і інших деталей будови. Одну з кожної пари гомологічних хромосом вносить в зиготу материнська, іншу - батьківська гамета.
Анафаза починається в момент поділу центромер, що утримували до цього разом обидві хроматиди кожної хромосоми. Все центромери діляться одночасно. Хроматиди, що стали тепер дочірніми хромосомами, роз'єднуються і розходяться до протилежних полюсів по нитках веретена, фіксованим у полюсів мітозу.
Телофаза - заключна стадія мітозу - починається завершенням руху хромосом до полюсів. Слідом за цим відбувається реконструкція інтерфазних ядер, яка як би повторює хід профази в зворотному порядку: формується ядерна мембрана, знову з'являються ядерця. Хромосоми, які тепер складаються з однієї нитки кожна, стоншуються, подовжуються і стають невидимими. В цей же час відбувається цитокинез - поділ цитоплазми. У кожній тваринній клітині утворюється перетяжка, поступово розділяє клітку на дві. У рослин поділ клітин завершується формуванням фрагмопласта - нової клітинної стінки між дочірніми ядрами.
Якщо на мітотичний діляться клітини подіяти колхіцином - алкалоїдом рослинного походження, що блокує полімеризацію ниток веретена, то в результаті хромосоми втрачають здатність пересуватися в площину веретена в прометафазі і розходитися в анафазе. Подвоєні хромосоми залишаються разом в одній клітці.
Найбільш характерний процес, що лежить в основі циклічних змін хромосом, - цикл спирализации-деспіралізаціі хромонем, або хромосомних ниток. На стадії ранньої профази мітозу, коли помітні в світловий мікроскоп хромосоми максимально деспіралізованние, можна бачити, що хромонеми нерівномірна по довжині: вона має тонкі і більш потовщені ділянки, які називаються хромомер. Розташування хромомер для кожної хромосоми щодо постійно. Хромомери представляють собою ділянки щільнішою спирализации хромонем.
Представлений коротко загальний хід мітозу, характерний для клітин тварин і рослин. Деякі відмінності від цього процесу зустрічаються, наприклад, у грибів (почкующиеся дріжджі-сахароміцети), у яких весь мітоз проходь! ' всередині ядерної мембрани, що зберігається на всіх стадіях, а ахроматинового веретено формується всередині ядра.