| Головна |
| «« | ЗМІСТ | »» |
|---|
У тварин сприйняття різних хімічних речовин, що знаходяться в навколишньому середовищі, здійснюється за допомогою двох хімічних аналізаторів: нюхового і смакового. Нюховий аналізатор є дистантних аналізатором, т. Е. Сприймає хімічні речовини, джерело яких знаходиться на відстані (іноді досить значній) від тварини. Нюховий аналізатор виконує функцію сигнальної хеморецепции. Крім пошуку і вибору їжі, порятунку від ворогів нюх допомагає при орієнтації як на суші, так і у воді. Хорошим прикладом цього служить повернення лососевих риб в батьківські водойми: на заключних етапах міграції, при попаданні лососевих в річки, основним орієнтиром для них служить знайомий запах, джерело якого знаходиться вище за течією. Істотну роль в житті тваринного грає так зване хімічне спілкування. Воно здійснюється за допомогою особливих хімічних речовин - «феромонів», які виробляються і виділяються спеціальними залозами. До феромонам відносяться статеві залучають речовини, використовувані для ме чення своєї території. Багато тварин по запаху відшукують або розпізнають статевого партнера. Відомо, що запуск барана в стадо овець безпосередньо перед сезоном спарювання прискорює і синхронізує течку у них. Причина цього полягає в тому, що запах барана викликає у овець частіші викиди лютеїнізуючого гормону, що стимулює прискорений розвиток фолікулів і овуляцію. Подібне явище спостерігається і у свиней, у яких статеве дозрівання значно прискорюється при ранньому запуску в стадо кнура. Запах дитинча може істотно впливати на прояв материнського інстинкту, що дозволяє зробити висновок про існування нюхового імпринтингу як у матері, так і у дитинчати. Тварин з добре розвиненим нюхом називають макросматікі - це майже всі ссавці. Значно слабкіше розвинений нюх у людини і у птахів - це мікросматіков.
Периферична частина нюхового аналізатора знаходиться в носовій порожнині: в області верхнього носового ходу і в задній верхній частині носової перегородки. Нюхова область пофарбована в більш темний колір: від жовто-бурого до темно-коріч-
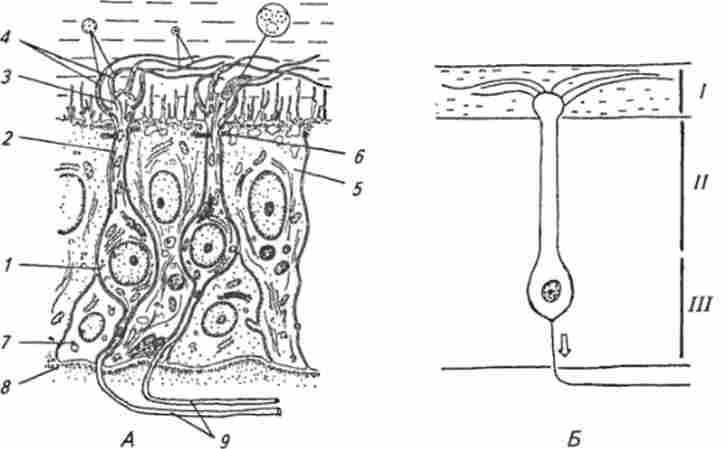
Мал. 15.10. ультраструктурная (А) і функціональна організація (Б) нюхового
рецептора:
/ - нюхова клітина; 2 - дендрит нюхової клітини; 3 булавовидное розширення дендрита; 4 нюхові волоски (показані поперечні зрізи волосків на різному видаленні від булавовидного розширення); 5-опорна клітина; 6 - зона щільного контакту; 7- базальна клітина; 8- базальнамембрана; 9- аферентні нервові волокна рецепторних клітин; / - область взаємодії пахучої речовини з рецепторними білками мембрани волосків і булавовидного розширення дендрита, де відбувається активація внутрішньоклітинних посередників (іАМФ, іГМФ, інозітолтріфосфат, іонів кальцію); // - область активації іонних каналів внутрішньоклітинними посередниками і генерація рецепторного потенціалу; /// - область трансформації рецепторного потенціалу в серію потенціалів дії невого на відміну від решти частини носової порожнини і являє собою епітеліальний шар товщиною 150 ... 200 мкм, що включає в себе нюхові, опорні і базальні клітини (рис. 15.10, А).
Нюхові клітини є біполярними нейронами з діаметром соми 5 ... 15 мкм (див. Рис. 15.10, Б). Від соми клітини до зовнішньої поверхні відходить дендрит діаметром 1 ... 2мкм і довжиною 5 ... 120 мкм. Вершина дендрита утворює булавовидное розширення діаметром 2 ... 3 мкм і виступає в шар слизу, що покриває нюхову область. Від нюхової «булави» відходять кілька тонких волосків - цілій, діаметр яких близько 100 ... 200 нм, а довжина 5 ... 250 мкм. Відростки нюхових клітин, що йдуть в ЦНС (аксони), належать до групи найбільш тонких нервових волокон, їх діаметр становить 0,2 ... 0,3 мкм. Число нюхових клітин у макросматіков може досягати сотень мільйонів.
Опорні клітини тісно примикають до нюхових клітин і утворюють у верхній частині зони щільний контакт (рис. 15.10, А), В результаті чого шар слизу відділяється від позаклітинної рідини, що оточує бічні і базальні ділянки клітин. На верхній (апікальної) поверхні опорних клітин є мікроволосків, але значно коротше, ніж у нюхових клітин. Базальні клітини лежать в глибині епітелію (див. Рис. 15.10, А) і становлять клітинний резерв, з якого утворюються рецепторні і опорні клітини.
Слизовий шар виступає в якості допоміжної структури у нюхових рецепторів і має велике значення для сприйняття хімічних речовин. Зокрема, слиз захищає нюховий епітелій від висихання у наземних тварин, а у водних - від зайвого змочування. Перш ніж станеться вплив молекул пахучої речовини на нюхові клітини, вони повинні розчинитися в слизу. Нерозчинні речовини не пахнуть. Зі слизом видаляються залишки пахучих молекул. Джерелом слизу є боуменовой залози, що знаходяться під епітеліальних шаром. Передбачається, що слиз можуть секретувати і опорні клітини.
При нормальному диханні тваринного через нюхову область проходить 5 ... 10% всього повітря, інша ж частина надходить в дихальну систему через нижній носовий хід. Слабкі запахи при такому диханні не сприймаються. Для того щоб пахнуть речовини досягли нюхових рецепторів, необхідно більш глибоке дихання або прискорене. Саме таким чином тварина принюхується, збільшуючи струм повітря через нюхову порожнину. Під час їжі нюхові рецептори дратуються повітрям, що проходить через хоани з ротової порожнини.
Малі розміри нюхових клітин істотно ускладнюють вивчення механізмів нюху на клітинному рівні, багато неясного і в питанні про адекватні подразники для нюхових клітин різних тварин. У навколишньому тварин повітрі знаходиться безліч хімічних сполук, які для одних тварин біологічно значимі і викликають відчуття, а для інших нейтральні і в даній концентрації не викликають збудження рецепторів і не створюють в ЦНС ніяких відчуттів. Тому при аналізі роботи нюхових клітин необхідно враховувати даний фактор.
Завдяки комплексним біохімічним, біофізичними та електро- трофізіологічним досліджень з'ясували процес сприйняття хімічного стимулу в нюхових рецепторах (див. Рис. 15.1). За відсутності стимуляції нюхові клітини генерують спонтанні аферентні потенціали дії, що вказує на де- поляризованность мембрани клітин. Вимірювання показали, що мембранний потенціал нюхових клітин на 30 ... 50 мВ менше за абсолютною величиною, ніж рівноважний калієвий потенціал (100 мВ). Таким чином, клітинна мембрана нюхових клітин в спокої проникна і для інших іонів (див. Гл. 2). Це значно збільшує чутливість рецептора до дії дратівної стимулу. При вдиханні пахучої речовини нюхові рецептори відповідають зміною частоти розрядів аферентних потенціалів дії - збільшенням або зменшенням, в залежності від якості і інтенсивності хімічного стимулу, але зміна її невелика. Наприклад, для найбільш досліджених нюхових клітин жаби вона рідко перевищує 10 імп / с. Зміни частоти потенціалів дії в аксонах нюхових клітин передує зміна мембранного потенціалу рецепторной клітини - рецепторного потенціалу. В одних випадках це деполяризация, при якій відповідно відбувається збільшення частоти аферентних потенціалів дії в аксоні, в інших - мембрана гиперполяризуется, що супроводжується зменшенням частоти спонтанних потенціалів дії або їх зникненням на час дії стимулу. Тривалість рецепторного потенціалу, а отже, і тривалість послідовності потенціалів дії у багато разів перевищує тривалість хімічного стимулу. По всій видимості, це обумовлено поступовим видаленням молекул пахучої речовини з нюхової порожнини.
Руйнування волосків значно знижує реакцію нюхових клітин на хімічні стимули. Це дозволило зробити висновок про те, що акт рецепції починається з дії молекули пахучої речовини на спеціальні ділянки мембрани - рецепторні білки волосків і булавовидний частини дендрита нюхової клітини. Детальні дослідження, проведені в кінці XX ст. на окремих нюхових клітинах різних тварин, виявили таку особливість в передачі хімічного стимулу: розташування рецепторних білків і іонних каналів, за рахунок активації яких генерується рецепторний потенціал, просторово рознесені; рецепторні білки знаходяться в мембрані апікальної частини дендрита - булавовидними утолщении і волосках, а іонні канали - в більш дистальних ділянках мембрани (див. рис. 15.10, Б). Активація іонних каналів відбувається через каскад реакцій за допомогою внутрішньоклітинних посередників. Послідовність подій наступна. Зовнішня сигнальна молекула зв'язується зі специфічним білком-рецептором в мембрані і активує білок-перетворювач, що передає сигнал через мембрану до ферменту-підсилювача, який, в свою чергу, активує внутрішньоклітинний посередник, який переносить сигнал всередині клітини. У якості посередників в нюхової клітці використовуються наступні речовини: циклічний аденозінмоно фосфат (цАМФ), циклічний гуанозинмонофосфат (цГМФ), інозітолтріфосфат, іони кальцію. Внутрішньоклітинні посередники можуть активувати різні іонні канали мембрани дендрита. Активація хлорних каналів викликає деполяризацію мембрани, що призводить до підвищення частоти потенціалів дії в аксоні. У той же час активація калієвих каналів зрушує мембранний потенціал в сторону гіперполяризації і пригнічує спонтанну активність.
Відповіді рецепторів на різні запахи до деякої міри індивідуальні та специфічні. При цьому у кожного рецептора досить широкий «спектр» чутливості на багато пахучі речовини. Розташовані поруч рецептори можуть мати зовсім різним характером специфічної чутливості. Початкові надії виділити порівняно невелику кількість функціональних типів нюхових рецепторів не виправдалися. Разом з тим здатність нюхового аналізатора до тонкої диференціації величезного числа запахів, а тим більше до аналізу багатокомпонентних сумішей передбачає існування досить великого числа паралельних шляхів, за якими передається інформація про запахи. Такі шляхи, ймовірно, обумовлені існуванням функціональних груп рецепторів, яких повинно бути досить багато. З огляду на, що у тварин-макросматіков число нюхових рецепторів може досягати декількох мільйонів, можна допустити у них наявність десятків або навіть сотень специфічних типів нюхових рецепторів.
Відходять від нюхових клітин аксони об'єднуються в пучки, кожен з яких оточений однією шванівської кліткою. Місцем закінчення аксонів є парні ділянки мозку, які мають характерну форму і отримали назву нюхових цибулин. Входження нюхових нервів в нюхові цибулини ипсилатерально, т. Е. Нерви не роблять перехрещення. Від нюхових цибулин ипсилатерально відходять кілька нервових стовбурів, які закінчуються в різних відділах переднього мозку: переднє нюхові ядро, нюховий горбок, пре- піріформная кора і періамігдалярная кора з прилеглою до неї кортікомедіальной частиною мигдалеподібного комплексу. Необхідно відзначити, що, мабуть, основна обробка інформації про нюховому стимулі здійснюється в нюхових цибулинах, оскільки вони є єдиним відділом мозку, двостороннє видалення якого завжди супроводжується повною втратою нюху.