| Головна |
| «« | ЗМІСТ | »» |
|---|
Смаковий аналізатор повідомляє тварині про хімічні властивості корму, що надходить в його ротову порожнину. Смакові рецептори на відміну від нюхових згруповані в спеціальні освіти, які за формою нагадують цибулини, - смакові цибулини. У свою чергу, переважна кількість смакових цибулин зібрані в особливих виростах епітелію мови - з-
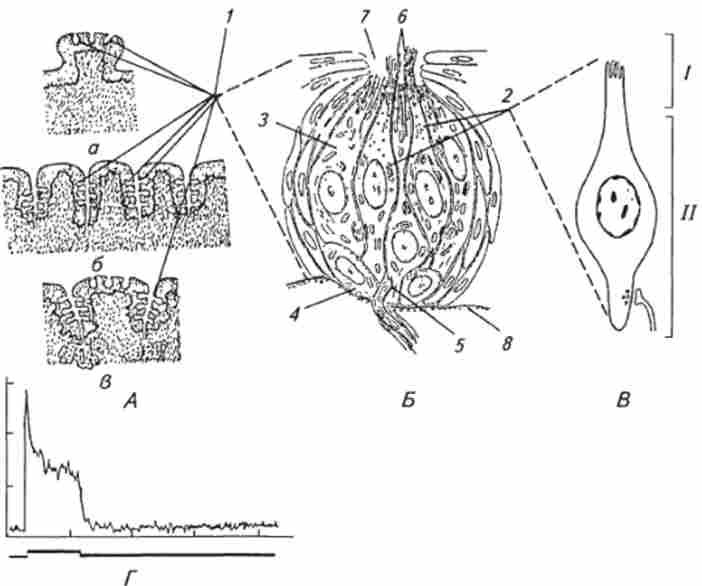
Мал. 15.11. Структурна і функціональна організація смакового рецептора:
А. а - грибоподібний сосочок; б - листоподібні сосочки; в - желобовідних сосочок; прямими лініями вказано місце розташування смакових цибулин (У) в сосочках. Б. Схема ультраструк- турной організації смакової цибулини мови кролика: 1 смакова цибулина; 2 рецепторная клітина, 3 - опорна клітина; 4-базальна клітина; 5-аферентних синапс; б-микроворсинки рецепторной клітини; 7-пора; 8- базальнамембрана. В. Рецепторная клітина: / - апикальная область рецепторной клітини, де відбувається взаємодія смакових всшсств з білками мембрани мікроворсинок. На апікальній мембрані іонні канали пов'язані з рецепцією кислих і солоних речовин; // - базолатсральная область рецепторной мембрани. Іонні канали пов'язані з рецепцією солодких речовин, які активуються внутрішньоклітинним посередником - цАМФ; Г. Зміна частоти потенціалів дії афферентного нервового стовбура (барабанної струни) при дії на смакові цибулини 0,1 м розчину хлориду натрію (солоний смак) По осі ординат - частота, імп / с, по осі абсцис - час, с; внизу- час дії сольового розчину Юс Сочка (рис. 15.11,> 4). Сосочки мають різну форму, і їх підрозділяють на грибоподібні, желобовідних і листоподібні. грибоподібний сосочки виступають над поверхнею мови і нагадують за формою гриб; розташовані в основному на передній дорсальній поверхні язика. листоподібні сосочки складаються з декількох вертикальних складок, розташованих паралельно, у вигляді листочків; сконцентровані по обидва боки задньої поверхні язика. желобовідних сосочки занурені в толше слизової мови та відокремлені від неї кільцеподібним желобком; локалізовані на дорсальній частині біля кореня язика. Рідкісні смакові цибулини знайдені також в слизовій оболонці задньої поверхні ротової порожнини, на стінках глотки, мигдаликів, надгортанника і гортані.
Смакова цибулина (див. Рис. 15.11, />) розташована в товщі багатошарового епітелію і орієнтована перпендикулярно верствам епітелію так, що апікальний відділ кожної цибулини сполучається з навколишнім середовищем, а базальна частина межує з сполучною тканиною, відокремленої базальноїмембраною. До складу смакової цибулини входять кілька типів клітин: смакові рецепторні клітини, опорні клітини і базальні клітини. смакова клітина має веретеноподібну форму; діаметр в найширшій частині 6 ... 10мкм (див. рис. 15.11, В). Апікальний відросток довший і тонкий, базальний - короткий, іноді ділиться і формує синапси з аферентні волокнами. На вершині апикального відростка є мікроворсинки діаметром близько 0,2 і довжиною 4 мкм. Смакові рецепторні клітини у верхній частині утворюють щільні контакти з опорними клітинами так, що є бар'єр між зовнішньою і міжклітинної середовищем латерального і базального ділянок клітин. На верхній частині опорних клітин мікроворсинки, за розмірами подібні до микроворсинками рецепторних клітин. Базальні клітини знаходяться в нижній частині смакової цибулини і формують щільні контакти з опорними клітинами. Верхня частина цибулини повідомляється із зовнішнім середовищем через вузький канал - пору. Під смакових цибулинах йде постійний процес оновлення клітин. Клітини безперервно діляться ми- тотічно і дегенерують, замінюючись новими. Так, смакова клітина в середньому живе (250 ± 50) ч. Молоді смакові клітини рухаються від периферії до центру, замінюючи старі клітини. Крім аферентних нервових волокон до смакових цибулин підходять і еферентні нервові волокна, утворюючи відповідно на базальних ділянках смакових клітин еферентні синапси (ланка зворотного зв'язку). Більшість нервових волокон, що утворюють синаптичні контакти з рецепторними клітинами смакових цибулин, локалізованих в передніх двох третинах мови, входять до складу язичного нерва. Нервові волокна, що іннервують смакові цибулини задньої третини мови, йдуть в ЦНС у складі язикоглоткового нерва. Невелика частина аферентних волокон від смакових клітин включається до складу інших черепномозкових нервів: трійчастого, лицьового і блукаючого. Аферентні волокна від смакових цибулин, що йдуть в складі язичного нерва, на деякій відстані виходять з нього і утворюють основну частину нервового стовбура, що отримав назву барабанної струни - horda tympani. Оскільки horda tympani складається головним чином з смакових аферентних нервових волокон, її використовують для електрофізіологічного дослідження функцій смакових рецепторних клітин.
При реєстрації електричної активності від нервових волокон horda tympani (див. Рис. 15.11, Г) було виявлено, що в них під час відсутності впливу подразника на смакові цибулини генеруються рідкісні потенціали дії. Це служить непрямим доказом того, що клітинна мембрана смакової клітини деполяризована і відбувається спонтанний викид медіатора з пресинаптичної області смакової клітини. Мікроелектродние вимірювання показали, що мембранний потенціал клітин зрушать на 40 ... 50 мВ в сторону деполяризації в порівнянні з рівноважним потенціалом для іонів калію, а це сприяє збільшенню чутливості рецептора.
Що ж служить адекватним подразником для смакових клітин мови? Для людини і тварин виділяють п'ять основних смакових відчуттів: солодке, гірке, кисле, солоне і смак їжі тваринного походження, обумовлений присутністю амінокислот, таких, як, наприклад, глютамат. Іноді це смакове відчуття називають японським терміном умами. Смакові відчуття виникають в тому випадку, якщо речовина розчиняється в слині і потім впливає на апикальную область смакових рецепторних клітин. На дію речовин, що володіють різним смаком, клітини реагують деполяризацією мембрани - генерацією рецепторного потенціалу, пов'язаної зі зміною іонної її провідності. Рецепторний потенціал збільшує амплітуду з підвищенням концентрації стимулюючого речовини; ця залежність носить логарифмічний характер. Незважаючи на труднощі з нанесенням дозованих хімічних стимулів на рецептор, вдалося показати, що у рецепторного потенціалу смакових клітин є динамічна і статична фази і відповідно швидка і повільна адаптації. Особливо це добре помітно при реєстрації аферентних потенціалів дії в нервових волокнах horda tympani (рис. 15.11, Г). Частота аферентних потенціалів дії максимальна на початку відповіді, далі йде спад, спочатку порівняно швидкий, а потім частота потенціалів дії повільно знижується до рівня, близького до початкового значення частоти спонтанної імпульсації. Безсумнівно, з адаптацією пов'язано поступове зниження і зникнення в деяких випадках відчуття смаку того чи іншого речовини.
З огляду на великі варіації в хімічній природі речовин, що викликають смакові відчуття, постає питання, за рахунок яких рецепторів апікальної мембрани і яким чином здійснюється рецепція специфічних речовин. Відповіді на ці питання були частково отримані в кінці XX ст. в дослідах на ізольованих смакових клітинах різних тварин. Виявилося, що рецепція хімічних речовин здійснюється декількома механізмами.
Відомо, що кислий смак пов'язаний з впливом на смакові рецептори кислот, причому ступінь «кіслості» залежить в основному від концентрації іонів водню. Дія кислот супроводжується виникненням деполяризационного потенціалу в смакових клітинах. Причиною деполяризації рецепторной мембрани є блокування іонами водню потенціалзавісімих калієвих каналів, розташованих на апікальній мембрані смакової клітини. Відповідно до рівняння Голдмана (див. Гл. 2) зменшення проникності для іонів калію зрушує мембранний потенціал в сторону деполяризації.
При дії на мову хлориду натрію виникає «класичний» солоний смак. Інші солі (по крайней мере для челозека) мають різні присмаки (гіркоти і т. Д.). Рецепція солоного пов'язана з наявністю на апікальній мембрані потснціалнеза- висимо іонних натрієвих каналів. Збільшення концентрації іонів натрію в слині сопровожается появою входить всередину клітини іонного натрієвого струму і деполяризацией мембрани.
Рецепцію речовин, що мають солодкий смак, забезпечують спеціальні рецепторні білки, що знаходяться на апікальній мембрані. Так само як і в нюхових клітинах іонні канали, за рахунок яких відбувається деполяризація мембрани, просторово відокремлені від «солодких» рецепторних білків. У передачі сигналу бере участь внутрішньоклітинний посередник - цАМФ, який знижує калиевую провідністьмембрани за допомогою блокування калієвих каналів, розташованих в базолатеральной мембрані клітини, на відміну від рецепції кислого.
Механізм рецепції гіркого і амінокислотного (умами) смаку розшифрований в меншій мірі. В апікальній мембрані смакових клітин є рецепторні білки для цих речовин: внутрішньоклітинним посередником для рецепції гіркого смаку виступають іони кальцію, а для амінокислот - цАМФ.
При проведенні експериментів на одиночних смакових клітинах з вивчення іонних каналів в різних ділянках мембрани була виявлена ще одна особливість. В базальних областях клітин є потенціалзалежні канали, за рахунок яких смакова клітина у відповідь на деполяризацію генерує потенціали дії. Таким чином, смакова клітина являє собою видозмінений первинний аферентних нейрон і послідовність подій в ній при рецепції хімічного стимулу збігається з такою в нюхових клітинах.
Поряд з смаковими клітинами, що реагують на одне специфічне смакове речовина, були виявлені клітини, які порушуються двома або трьома смаковими речовинами. Тварини часто по-іншому реагують на смакові речовини, ніж людина, суб'єктивні реакції якого використовуються для пояснення роботи смакових клітин. Так, неприємний для людини гіркий смак не викликає у деяких видів тварин негативного ставлення до нього: кролики і коні охоче поїдають гірку осиковий кору; кислий смак лимона дуже неприємний для коней, корів, собак.
Центральні відростки нейронів, що іннервують смакові цибулини, закінчуються в довгастому мозку в ядрі одиночного пучка, де відбувається перемикання на нейрони наступного порядку. Аксони цих нейронів утворюють частковий перехрест в ретикулярної формації і досягають вентромедиального комплексу ядер таламуса. З таламуса аксонинейронів третього порядку направляються в кору головного мозку в корковий центр смаку, локалізація якого поки точно не визначена. Чутливість смакових клітин контролює ЦНС за допомогою еферентної ланки зворотнього зв'язку за допомогою еферентних синапсів.
У тварин смакова чутливість змінюється в залежності від функціонального стану шлунково-кишкового тракту. Сигнали з рецепторів слизової шлунка йдуть в ЦНС, яка регулює збудливість смакових рецепторів. Розвиток смакового аналізатора у тварин визначається складом поедаемой їжі. Найбільшою мірою розвинений смаковий аналізатор у травоїдних тварин, так як їх їжа включає велику кількість трав різних видів, серед яких вони повинні вибирати їстівні рослини. У м'ясоїдних тварин їжа більш одноманітна: м'ясо не може принести їм шкоди, тому й не виникає необхідність тонко розрізняти його смак.