| Головна |
| «« | ЗМІСТ | »» |
|---|
Слуховий аналізатор призначений сприймати механічні коливання (звукові хвилі) із зовнішнього середовища, створювані джерелами, що знаходяться на відстані, іноді досить значному, від тварини. Звукопередающей середовищем для наземних тварин є повітря, а для водних тварин - вода, в якій звук поширюється в 4 рази швидше, ніж в повітрі. Звукові сигнали, що сприймаються тваринами, підрозділяють на звуки небіологічного походження (залежні від сили вітру, випадання опадів, шуму листя дерев та ін.) І звуки біологічного походження, пов'язані з сигналами акустичного спілкування різних тварин, а також шерехами і шумами, виробленими хижаками або їх жертвами.
Розвиток слухового аналізатора в процесі еволюції досягло найбільшого досконалості у ссавців. Периферична частина слухового аналізатора складається з трьох відділів: зовнішнього, середнього і внутрішнього вуха (рис. 15.12). Зовнішнє вухо включає в себе вушну раковину і зовнішній слуховий прохід. За своєю формою зовнішнє вухо нагадує воронку, яка направляє звукові хвилі в слуховий прохід, т. е. забезпечує покращене сприйняття звуків. У багатьох тварин рефлекторне управління рухливістю вушної раковини добре розвинене за допомогою спеціальних м'язів, що не вимагає повороту голови. Раз
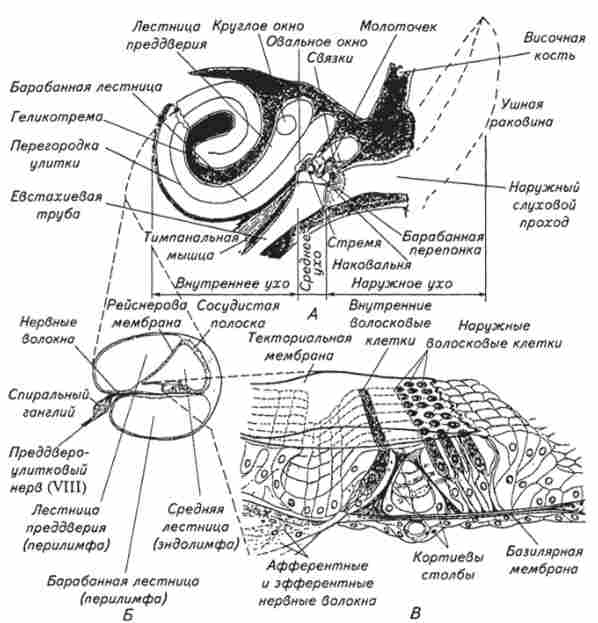
Мал. 15.12. Вухо ссавців:
А. Загальна схема будови (півкруглі канали не показані); Б. Поперечний розріз через равлика; В. Схема кортісвого органу
заходи і форма вушної раковини варіюють у тварин різних видів в значній мірі: дуже рухливі вушні раковини у кішок, коней, деяких порід собак.
Зовнішній слуховий прохід являє собою вузьку, кілька викривлену трубку, по якій звукові коливання проходять всередину вуха. Спеціальні залози в шкірі зовнішнього слухового проходу виробляють секрет - вушну сірку, яка охороняє вухо від забруднення і перешкоджає висиханню барабанної перетинки, яка відділяє зовнішнє вухо від середнього. Барабанна перетинка кріпиться на внутрішньому кістковому кінці зовнішнього слухового проходу (див. Рис. 15.12, /!), Товщина її становить
100 ... 200 мкм. Призначення барабанної перетинки полягає в передачі без спотворень частоти впливають на неї звукових коливань. Необхідною умовою для цього є відсутність у неї резонансних явищ, т. Е. Вона не посилює звукові коливання. Це можливо завдяки особливостям її будови, а також розташуванню її в слуховому проході. Барабанна перетинка - це сполучнотканинна мембрана, утворена з радіальних і кільцевих волокон. Ступінь натягу її окремих ділянок неоднакова; в звуковому проході вона розташована косо і середина її втягнута всередину.
Середнє вухо - це трехкосточковая звукопередающая важільна система, що включає в себе послідовно з'єднані кісточки: молоточок, ковадло і стремячко. Велика частина поверхні барабанної перетинки контактує з ручкою молоточка, який зчленовується зв'язками з ковадлом. Ковадло з'єднана зі стремечком, пов'язаним з мембраною овального віконця, що відкривається в порожнину передодня (внутрішнє вухо), заповненої рідиною. Розташування та механічні властивості сруктур середнього вуха забезпечують оптимальну передачу звукових коливань з повітряного середовища в рідку середу внутрішнього вуха. Зокрема, ефективна частина барабанної перетинки (т. Е. Що знаходиться в контакті з молоточком) приблизно в 17 разів більше, ніж площа підстави стремечка, крім того, спосіб обертання кісточок додатково збільшує тиск на овальне вікно - отвір, що веде у внутрішнє вухо. Завдяки всьому цьому тиск на овальне вікно в порівнянні з барабанною перетинкою збільшується приблизно в 20 разів. Таким чином, система важеля середнього вуха зменшує амплітуду переміщень барабанної перетинки, перетворюючи їх в незначні за розмахом, але в відповідне число раз посилених по тиску переміщень стремечка. Цей механізм збільшення тиску є надзвичайно важливим пристосуванням, що забезпечує ефективну передачу акустичної енергії з повітряного середовища в рідку.
В системі середнього вуха функціонують кілька механізмів, що оберігають пошкодження його структур від вельми сильних звукових коливань. Так, порожнину середнього вуха не зачинені, а з'єднується з порожниною глотки особливим каналом - евстахиевой трубою, служить для вирівнювання тиску по обидві сторони барабанної перетинки. Це має велике значення для запобігання барабанної перетинки при значній різниці тиску в барабанної порожнини і в навколишньому середовищі. Така різниця може виникнути при потрапляє в вухо звукової хвилі, наприклад від гуркоту сильного грому. Розкриття євстахієвої труби сприяє вирівнюванню тиску і попереджає розрив барабанної перетинки.
Інший охоронний механізм полягає в рефлекторному скороченні м'язів середнього вуха у відповідь на надмірно сильні звуки. Пов'язана з молоточком тімпанальная м'яз, скорочуючись, зміщує барабанну перетинку всередину, а одночасне скорочення м'язи стремечка обмежує рух стремечка і мембрани овального вікна, що в сукупності призводить до зменшення тиску, переданого важільним механізмом.
У кісткової перегородки, яка відділяє середнє вухо від внутрішнього, крім овального вікна є також кругле вікно, або вікно равлики. Воно затягнуто тонкою сполучнотканинною мембраною і може служити додатковим пристосуванням для передачі звукових коливань в разі пошкодження барабанної перетинки і кісточок.
Внутрішнє вухо розташоване в кам'янистій частині скроневої кістки і складається з кісткового і перепончатого лабіринтів (Див. Рис. 15.12, Б). До складу периферичної частини слухового аналізатора входить лише частина перепончатого лабіринту, який поміщається в кістковому, як у футлярі, і точно повторює його форму. Звуковий лабіринт є звужується до кінця трубку, спірально закручену на зразок раковини - равлики. Число витків равлики у різних ссавців буває різний: у людини - 2,5 витка, у кішки - 3, у свині - 4. Кістки равлик розділена вздовж кісткової спіральної стінкою, яка, однак, не доходить до протилежної стінки, а її продовжує основна - базилярная мембрана. На початку равлики кісткова спіральна стінка має найбільшу ширину, а базилярная мембрана найменшу (див. Рис. 15.12, Б). У міру руху до вершини равлики ширина кісткової стінки зменшується, а базилярної мембрани збільшується. Комбінація кісткової перегородки з базилярної мембраною ділить просвіт равлики на два головних поздовжніх канали: барабанна драбина і сходи передодня. Вони заповнені рідиною, званої перилимфой, і повідомляються між собою у верхівки равлики через невеликий отвір - гелікотрему, діаметром близько 20мкм. Між цими каналами лежить третій канал - внутрішні сходи, відокремлений від них базилярної і рейснеровой мембраною, заповнений рідиною - ендолімфою. Перилимфа і ендолімфа у ссавців відрізняються в значній мірі за іонним складом один від одного, а також від плазми крові. Так, концентрація іонів калію в ендолімфі в
30 ... 35 разів перевищує їх концентрацію в плазмі крові, а в пери лимфе - приблизно в 1,5 ... 2,5 рази. І навпаки, концентрація натрію в ендолімфі приблизно в такій же пропорції нижче, ніж в плазмі крові і перилимфе.
Причина іонної асиметрії пов'язана з функціонуванням електрогенних іонних насосів. Різниця в концентрації заряджених частинок супроводжується виникненням різниці потенціалів. У каналах равлики виявили позитивні (щодо плазми крові) ендокохлеарние і ендовестібуляр- ні потенціали. Роль їх в роботі слухового аналізатора поки не з'ясована.
На базилярній мембрані розташовується рецептор звукового аналізатора - кортів орган (Див. Рис. 15.12, Я), що складається з чотирьох рядів волоскових клітин - одного внутрішнього і трьох зовнішніх. На верхній частині рецепторних клітин кортиевого органу перебуває кілька десятків волосків - стереоцилій. Нижче рейснеровой мембрани (базилярної) від кісткової стінки равлики відходить текторал'ная (Покривна) мембрана, нависає над рядами розташованих під нею волоскових клітин. Стереоціліямі волоскових клітин трьох зовнішніх рядів контактують з текторальной мембраною, а внутрішнього ряду з нею контакту не мають.
Всі згадувані вище структури є структури допоміжного апарату (рис. 15.13). Вони призначені передавати з мінімальними спотвореннями звукові коливання до рецепторних волоскові клітини. Волоскові клітини мають довжину 20 ... 40 мкм і діаметр 5 ... 10мкм. Стереоціліямі на апікальній частині рецепторних клітин мають довжину 4 ... 10мкм і діаметр 0,1 ... 0,3 мкм. Серцевину стереоцилій утворює пучок фібрил, які зверху оточені клітинною мембраною. У підстави стереоцилій фібрили зливаються і закінчуються всередині клітини в кутікулярной платівці. Стереоціліямі у волоскових клітин трьох зовнішніх рядів розташовуються у вигляді підковоподібної фігури, причому найдовші знаходяться на вершині, а внутрішнього ряду - на апікальній поверхні в кілька рядів. Крім волоскових клітин в Кортиєва органі є кілька типів опорних клітин. Первинний аферентних нейрон контактує з волосовими клітинами за допомогою синаптичних закінчень. До рецепторних клітин підходять і еферентні нервові волокна, утворюючи відповідно еферентні синапси (ланка зворотного зв'язку).
Аферентні волокна слухового нерва генерують спонтанні потенціали дії, і частота їх іноді буває досить високою (більше 10 імп / с). Наявність спонтанної імпульсації зв'язується з фоновим виділенням медіатора в синаптичній області волоскових клітин, що, в свою чергу, передбачає деполяризацию мембрани волоскових клітин. Мікроелектродние вимірювання показали, що дійсно мембранний потенціал волоскових клітин зрушать на
40 ... 60 мВ в деполяризационного сторону від рівноважного калієвого потенціалу, що може бути однією з причин високої чутливості слухового аналізатора.
Коливання рідини в равлику, викликані стимуляцією стремечком мембрани овального вікна, передаються базилярної мемб-
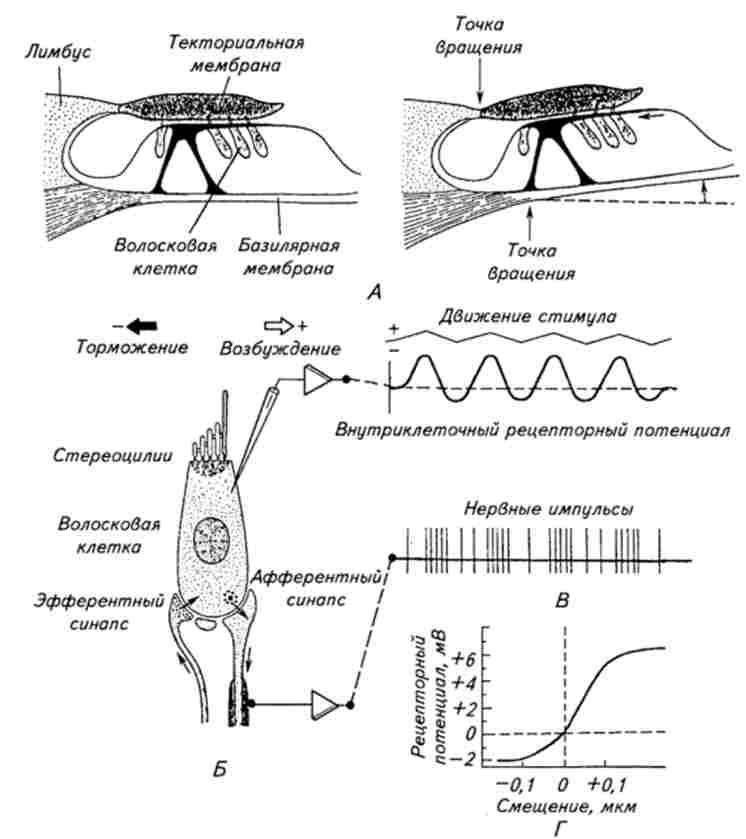
Мал. 15.13. Схема збудження волоскових клітин і генерації в них електричної
активності:
А Виникнення зрушує сили, що впливає на стереоціліямі рецепторних клітин при відхиленні базилярної мембрани вгору. Ковзання тскторальной мембрани по кортіс- вому органу обумовлено тим, що тскторальная і базилярная мембрани мають різні точки обертання. Переміщення мембрани дані в сильно збільшеному масштабі; Б Волосковая рецепторная клітина і її електрична активність при механічному подразненні; В. Рецепторний потенціал і аферентна імпульсна активність при різному напрямку механічного роздратування, що впливає на стереоціліямі; Г Залежність між амплітудою зміщення стсреоіілій і величиною рецепторного потенціалу рані, на якій покоїться кортів орган. При цьому відбувається механічне зміщення волосків на апикальном кінці рецепторной клітини, активація механоактівіруемих каналів і виникнення рецепторного потенціалу. Вважають, що зрушення стереоцилій обумовлений різним розташуванням центрів обертання базилярної мембрани і текторальной мембрани, до якої прикріплені зовнішні волоскові клітини (див. Рис. 15.13, /!). Завдяки такому розташуванню відхилення базилярної мембрани вгору викликає зміщення кінчиків стереоцилій в напрямку їх вершини. При цьому відбувається посилення натягу мембрани і активація механоактівіруемих іонних каналів. І навпаки, зміщення базилярної мембрани вниз викличе нахил стереоцилій в протилежному напрямку, що зменшить натяг рецепторной мембрани. Про це свідчать дані, отримані при вивченні одиночних волоскових клітин (див. Рис. 15.13, В). Зсув пучка стереоцилій в сторону найбільш довгих стереоцилій, що знаходяться на вершині подковообразного розташування стереоцилій, викликає деполяризацію мембрани рецепторної клітини. Зсув же стереоцилій в іншу сторону гіперполяризуючий мембрану (див. Рис. 15.13, В). Таким чином, в спокої частина механоактівіруемих каналів відкрита і ослаблення натягу мембрани викликає їх інактивацію і зменшення іонного струму. На ізольованих волоскових клітинах за допомогою методики локальної фіксації було вивчено дію окремих механоактівіруемих каналів. Реакція механоактівіруемих каналів слухових волоскових клітин на механічне подразнення, як і інших іонних каналів, являє собою ступенчатообразние коливання струму різної тривалості, але однаковою амплітуди. Залежність сумарного іонного струму волоскових клітин від амплітуди механічного стимулу носить логарифмічний характер (див. Рис. 15.13, Г).
Інший етап роботи периферичної ланки слухового аналізатора полягає в кодуванні інформації про висоту (частоті) звуку. Тварини здатні сприймати звуки з частотою
15 ... 80000 Гц. При частоті звуку, що перевищує 1000 Гц, однозначної відповідності між звуковими коливаннями і частотою аферентних потенціалів дії не може бути, що обумовлено наявністю абсолютного рефрактерного періоду після кожного потенціалу дії. Тому інформація про частоту звуку повинна кодуватися НЕ частотою потенціалів дії, а за допомогою іншого механізму. Вивчаючи морфологічні та механічні властивості базилярної мембрани, Г. Бекеш в 60-х роках XX ст. прийшов до висновку, що амплітуда зміщення різних її ділянок залежить від частоти звуку. Причому коливання в базилярної мембрани поширюються в вигляді біжучої хвилі. Поступове збільшення механічної податливості базилярної мембрани від її вузького кінця до широкого в дистальної частини равлики призводить до того, що амплітуда біжать по ній хвиль змінюється (рис. 15.14).
Точка, в якій зміщення мембрани має найбільшу амплітуду (і відповідно відбувається максимальна стимулівМал. 15.14. Амплітуда зміщення базилярної мембрани в залежності від звукових частот:
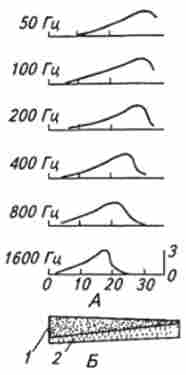
А. Амплітуда зміщення базилярної мембрани дана в відносних одиницях; по осі абсцис - відстань уздовж мембрани від драбина, мм; Б. Перегородка равлики: 1 кісткова спіральна стінка, звужується до дистальному кінця; 2 базилярная мембрана, що розширюється до дистальному кінця
ція стереоцилій волоскових клітин), залежить від частоти хвиль, що біжать, а отже, і від частоти звуку. При впливі високих частот біжать хвилі викликають максимальне зміщення базилярної мембрани ближче до овального вікна. Для низькочастотних сигналів область найбільшого зміщення базилярної мембрани зсувається до дальнього кінця равлики. Таким чином, принцип кодування частоти звуку - просторовий. У той же час кодування гучності (амплітуди) звукового стимулу здійснюється частотним кодом. Ступінь зсуву базилярної мембрани в будь-якій точці її довжини визначає силу стимуляції волоскових клітин і тим самим амплітуду рецепторного потенціалу, що, в свою чергу, обумовлює частоту імпульсів в аферентних нервових волокнах, що іннервують дану групу клітин. У волоскових клітин, як і в інших Механорецептори, у відповідь на тривалу дію механічного стимулу спостерігається адаптація рецепторного потенціалу. Це викликає зменшення викиду медіатора і афферентной імпульсації. Дане явище лежить в основі адаптації слуху, т. Е. Тимчасове зниження чутливості слухового аналізатора при тривалій дії звукового сигналу.
Першими аферентні нейронами слуховий системи ссавців є клітини спірального ганглія равлики. Периферичні відростки цих клітин направляються у вигляді спіральних і радіальних пучків до рецепторних клітин кортіе- вого органу і утворюють на них синапси. Аксони цих нейронів формують завитковому, або кохлеарную, гілка слухового нерва, який, пройшовши через внутрішній слуховий прохід, входить в мозковий стовбур і закінчується в кохлеарних ядрах мозку. Улітку проектується на кохлеарні ядра таким чином, що волокна від базального високочастотного завитка равлики закінчуються в дорсомедіальних частини кохлеарних ядер, а від апікального - низькочастотного - в вентролатеральной частини. Тіла аферентних нейронів третього порядку знаходяться в задніх буграх четверохолмия і в медіальному колінчастому тілі таламичної області. Внутрішнє коленчатое тіло - це таламичний центр слухової системи. Аксони клітин внутрішнього колінчастого тіла закінчуються на клітинах слуховий області кори - кінцевого етапу слухового шляху, розташованого в скроневій частці великих півкуль. Завдяки наявності двох вух тварини точно визначають напрямок, звідки доноситься звук. Це явище отримало назву бинаурального ефекту. У скроневих частках кори відбувається аналіз звукових коливань і створюється уявлення про спрямованість звуку. При двосторонньому руйнуванні слуховий кори просторовий слух порушується.