| Головна |
| «« | ЗМІСТ | »» |
|---|
Раніше були розглянуті характерні риси фенотипічного прояву і успадкування окремих ознак. Однак фенотип організму являє собою сукупність багатьох властивостей, за формування яких відповідають різні гени. Так як загальне число генів у генотипі значно більше числа хромосом, кожна хромосома містить в собі комплекс генів. У зв'язку з цим неалельні гени можуть розташовуватися або в різних хромосомах, або входити до складу однієї з них, займаючи різні локуси АБС, КМР. Цим визначається характер успадкування групи ознак, яке може бути незалежним або зчепленим.
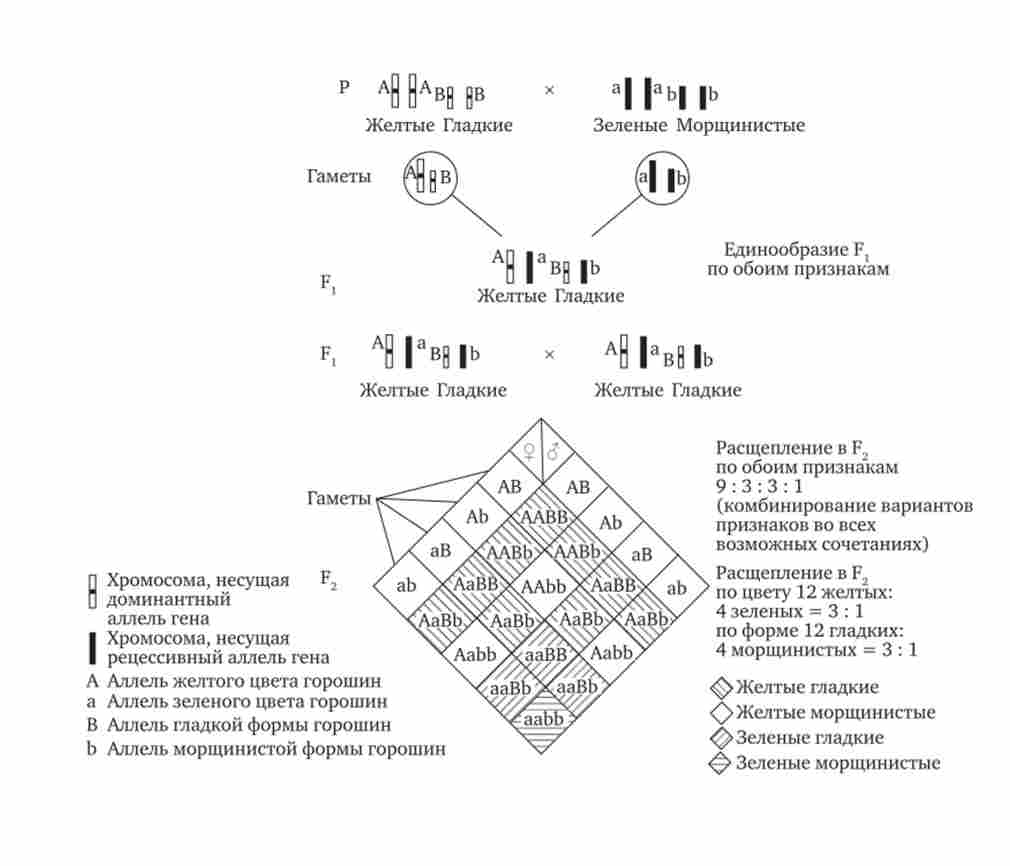
Незалежне успадкування ознак вперше було описано Г. Менделем в дослідах на горосі, коли одночасно аналізувався спадкування в ряду поколінь кількох ознак, наприклад кольору і форми горошин (рис. 7.11). Кожен з них окремо підкорявся закону розщеплення в F2. В Водночас різні варіанти цих ознак вільно комбінували у нащадків, зустрічаючись як в поєднаннях, які спостерігаються у їхніх батьків (жовтий колір і гладка форма або зелений колір і зморшкувата форма), так і в нових поєднаннях (жовтий колір і зморшкувата форма або зелений колір і гладка форма). На підставі аналізу отриманих результатів Г. Мендель сформулював закон незалежного успадкування ознак, відповідно до якого: «Різні пари ознак, що визначаються неалельних генами, передаються нащадкам незалежно один від одного і комбінуються у них у всіх можливих поєднаннях».
Цьому закону підкоряються в першу чергу неалельні гени, розташовані в негомологічних хромосомах. У мейозі ці хромосоми утворюють зі своїми гомологами різні пари, або біваленти, які в метафазі I мейозу випадково шикуються в площині екватора веретена поділу. Потім в анафазе I мейозу гомологи кожної пари розходяться до різних полюсів веретена незалежно від інших пар. В результаті у кожного з полюсів виникають випадкові поєднання батьківських і материнських хромосом в гаплоидном наборі (див. Рис. 3.75). Отже, різні гамети містять різні комбінації батьківських і материнських алелей неалельних генів.
Різноманітність типів гамет, утворених організмом, визначається ступенем його гетерозиготности і виражається формулою 2 ", де
Мал. 7.11.
Незалежне успадкування ознак (кольору і форми горошин)
290 Глава 7. Онтогенез як процес реалізації спадкової інформації
п - число локусів в гетерозиготному стані. У зв'язку з цим діге- терозіготние гібриди F, утворюють чотири типи гамет з однаковою ймовірністю. Реалізація всіх можливих зустрічей цих гамет при заплідненні призводить до появи в F2 чотирьох фенотипічних груп нащадків у співвідношенні 9: 3: 3: 1. Аналіз нащадків F2 по кожній парі альтернативних ознак окремо виявляє розщеплення в співвідношенні 3: 1.
Відкриття незалежного характеру успадкування різних ознак у гороху дало можливість Г. Менделя висловити припущення про дискретності спадкового матеріалу, в якому за кожну ознаку відповідає своя пара спадкових задатків, які зберігали в ряду поколінь свою структуру і не змішуються один з одним. Сучасні уявлення про надмолекулярної організації спадкового матеріалу в хромосомах і закономірності їх передачі в ряду поколінь клітин і організмів пояснюють незалежний характер успадкування ознак розташуванням відповідних генів в негомологічних хромосомах.
У дослідах Г. Менделя спадкова конституція гібридів F: встановлювалася на основі аналізу результатів їх самозапилення, одержуваних в F2. В даний час з метою з'ясування генотипу організмів з домінантним фенотипом (гомо- або гетерозигота) широко застосовують також аналізує схрещування. Воно полягає в схрещуванні організму, генотип якого необхідно визначити, з організмом, що несе рецесивний ознака, а отже, гомозиготних по рецесивним аллели (рис. 7.12).
Так як гомозиготні організми утворюють один тип гамет: аа - (Т), aabb - (ab), aabbcc - (abc) і т. Д., - при анализирующем схрещуванні кількість фенотипів нащадків залежить від числа типів гамет, утворених організмом з домінантним фенотипом . Якщо останній гомозиготен по аналізованих генам, то він також утворює тільки один тип гамет і потомство від аналізує схрещування одноманітно і має домінантний фенотип (рис. 7.12,1).
Якщо аналізований організм гетерозиготний по одному гену, він утворює два типи гамет і при анализирующем схрещуванні з'являються нащадки двох різних фенотипів з домінантним або рецесивним ознакою (рис. 7.12, II).
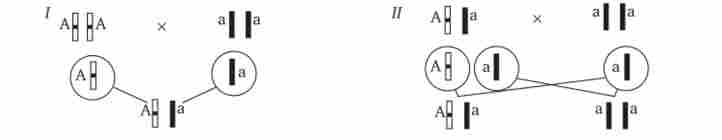
Мал. 7.12. Аналізує (моногибридное) схрещування. Пояснення см. В тексті
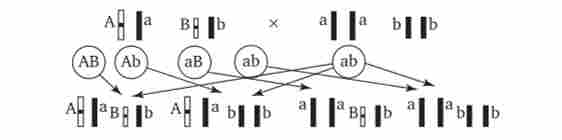
Дігетерозіготний організм при анализирующем схрещуванні дає чотири види нащадків (рис. 7.13).
Мал. 7.13. Аналізує (дигибридное) схрещування. Пояснення см. В тексті
У тому випадку, коли неалельні гени А і В успадковуються незалежно, розташовуючись в різних хромосомах, дігетерозіготний організм утворює чотири типи гамет з однаковою ймовірністю. Тому в результаті аналізує схрещування чотири фенотипично розрізняються виду нащадків з'являються в співвідношенні 1: 1: 1: 1 і несуть різні поєднання варіантів двох ознак.
Аналіз успадкування одночасно декількох ознак у дрозофіли, прознавств Т. Морганом, показав, що результати аналізує схрещування гібридів Fj іноді відрізняються від очікуваних при незалежному успадкуванні. У нащадків такого схрещування замість вільного комбінування ознак різних пар спостерігали тенденцію до спадкоємства переважно батьківських поєднань ознак. Таке спадкування ознак було названо зчепленим. Зчеплене успадкування пояснюється розташуванням відповідних генів в одній і тій же хромосомі. У складі останньої вони передаються з покоління в покоління клітин і організмів, зберігаючи поєднання алелей батьків.
На рис. 7.14 представлені результати аналізу успадкування забарвлення тіла і форми крил у дрозофіли, а також їх цитологічне обґрунтування. Звертає увагу, що при анализирующем схрещуванні самців з Fj з'являлося всього два види нащадків, схожих з батьками по поєднанню варіантів аналізованих ознак (сіре забарвлення тіла і нормальні крила або чорне забарвлення тіла і короткі крила) в співвідношенні 1: 1. Це вказує на освіту самцями Fj всього двох типів гамет з однаковою ймовірністю, в які потрапляють вихідні батьківські поєднання алелей генів, що контролюють названі ознаки (BV або bv).
При анализирующем схрещуванні самок Fa з'являлося чотири види нащадків з усіма можливими поєднаннями ознак. При цьому нащадки з батьківськими поєднаннями ознак зустрічалися в 83%. У 17% нащадків були нові комбінації ознак (сіре забарвлення тіла і короткі крила або чорне забарвлення тіла і нормальні крила). Видно, що і в цих схрещуваннях проявляється схильність до зчеплення спадкоємства або домінантних ознак, або рецесивних (83%). часткове порушення
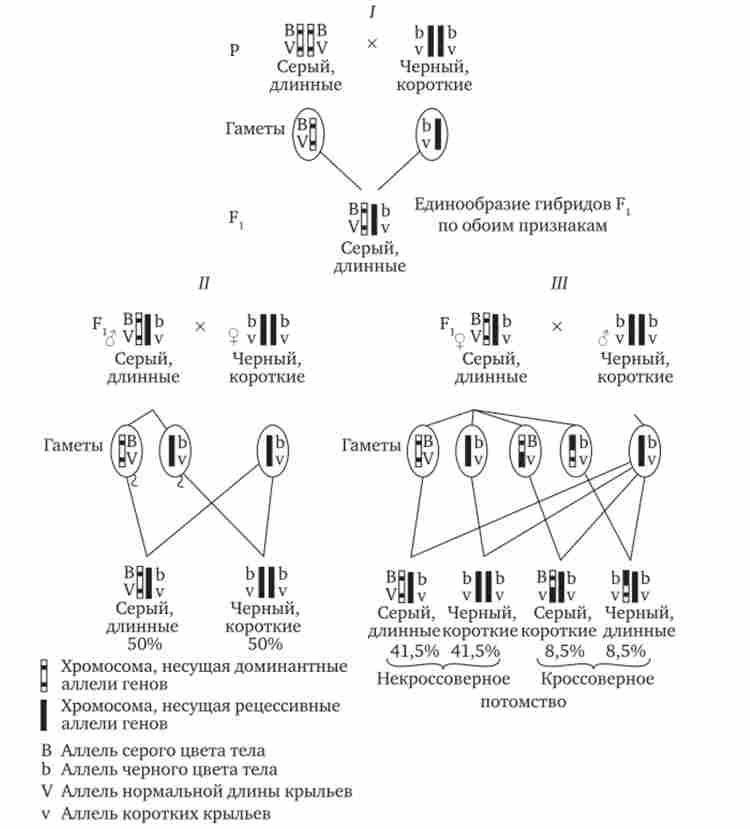
Мал. 7.14. Зчеплене успадкування ознак (кольору тіла і довжини крил у дрозофіли).
I-схрещування чистих ліній; II, 111 - аналізує схрещувань самців і самок з Fj
зчеплення (17% нащадків) було пояснено процесом кроссінго- віра - обміном відповідними ділянками гомологічних хромосом в профазі I мейозу (див. рис. 3.72).
З результатів схрещування слід, що самки дрозофіли утворюють чотири типи гамет, більшість з яких (83%) некроссоверние ((в?) І (bv)), 17% утворених ними гамет з'являються в результаті кросинговеру і несуть нові комбінації алелей аналізованих генів ((Bv) або (bv)). Відмінності, що спостерігаються при схрещуванні самців і самок з F1 з рецесивними гомозиготними партнерами пояснюються тим, що з малозрозумілих причин у самців дрозофіли не відбувається кросинговеру. В результаті самці-дигетерозиготи по генам, розташованим в одній хромосомі, утворюють два типи гамет. У самок кроссинговер має місце і призводить до утворення некроссоверних і кроссоверних гамет, по два типу кожних. Тому в потомстві від аналізує схрещування з'являється чотири фенотипу, два з яких мають новими в порівнянні з батьками поєднаннями ознак.
Вивчення успадкування інших поєднань ознак показало, що відсоток кроссоверного потомства для кожної пари ознак завжди один і той же, але він розрізняється для різних пар. Це спостереження стало підставою для укладення, що гени в хромосомах розташовані в лінійному порядку. Вище зазначалося, що хромосома є групою зчеплення певних генів. Гомологічні хромосоми - це однакові групи зчеплення, які відрізняються один від одного лише алелями окремих генів. При кон'югації гомологи зближуються своїми аллельними генами, а при кроссін- Говера вони обмінюються відповідними ділянками. В результаті з'являються кроссоверние хромосоми з новим набором алелей. Частота, з якою відбувається обмін на ділянці між двома даними генами, залежить від відстані між ними (правило Т. Моргана). Відсоток кроссоверних гамет, що несуть кроссоверние хромосоми, побічно відображає відстань між генами. Це відстань прийнято виражати в сантіморганідах. За одну сантіморга- Ніду приймають відстань між генами, при якому утворюється 1% кроссоверних гамет.
При збільшенні відстані між генами збільшується ймовірність кросинговеру на ділянці між ними в клітинах - попередницях гамет. Так як в акті кросинговеру беруть участь дві хро- матіди з чотирьох, присутніх в біваленте, то навіть у разі здійснення обміну між генами даної пари у всіх клітинах - попередницях гамет відсоток кроссоверних статевих клітин не може перевищити 50 (рис. 7.15). Однак така ситуація можлива лише теоретично. Практично зі збільшенням відстані між генами зростає можливість проходження одночасно декількох кроссинговеров на даній ділянці (див. Рис. 5.9). Так як кожен другий перехрест призводить до відновлення колишнього поєднання алелей в хромосомі, зі збільшенням відстані число кроссоверних гамет може не збільшуватися, а зменшуватися. З цього випливає, що відсоток кроссоверних гамет є показником істинного відстані між генами лише при досить близькому їх розташуванні, коли можливість другого кросинговеру виключається.
Порушення зчепленого успадкування батьківських алелей в результаті кросинговеру дозволяє говорити про неповному зчепленні на відміну від повного зчеплення, спостережуваного, наприклад, у самців дрозофіли.
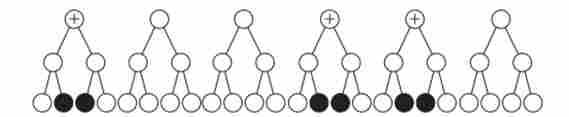
Мал. 7.15. Схема, яка пояснює низький відсоток кроссоверних гамет (по відношенню до двох даними генам).
Плюсом позначені клітини - попередниці гамет, в яких кроссинговер пройшов на ділянці між двома даними генами; зачорнені кроссоверние гамети
Використання аналізує схрещування в дослідах Т. Моргана показало, що з його допомогою можна з'ясовувати не тільки склад пар неалельних генів, але і характер їх спільного успадкування. У разі зчепленого успадкування ознак за результатами аналізує схрещування можна встановити також відстань між генами в хромосомі.