| Головна |
| «« | ЗМІСТ | »» |
|---|
На характер успадкування в ряду поколінь складних ознак певний вплив має тип взаємодії неалельних генів (див. Розд. 3.6.5.2). Різні комбінації їх алелей можуть забезпечувати появу нового ознаки або його варіанти, зникнення ознаки, зміна характеру його прояви у нащадків.
Нижче будуть розглянуті закономірності успадкування ознак при незалежному успадкуванні взаємодіючих неалельних генів.
У тому випадку, коли складний ознака визначається декількома парами генів в генотипі і їх взаємодія зводиться до накопичення ефекту дії певних алелей цих генів, серед нащадків гетерозигот спостерігається різна ступінь вираженості ознаки, що залежить від сумарної дози відповідних алелей. Наприклад, ступінь пігментації шкіри у людини, яка визначається чотирма парами генів, коливається від максимально вираженою у гомозигот по домінантним аллелям у всіх чотирьох парах Д ° мінімальної у гомозигот по рецесивним аллелям (Р1Р1Р2Р2Р3Р3Р4Р4) (см- Рис* 3.80). При шлюбі двох мулатів, гетерозиготних по всіх чотирьох парах, які утворюють по 24 = 16 типів гамет, виходить потомство, 1/256 якого має максимальну пігментацію шкіри, 1/256 - мінімальну, а решта характеризуються проміжними показниками експресивності цієї ознаки. У розібраному прикладі домінантні аллели полигенов визначають синтез пігменту, а рецесивні - практично не забезпечують цієї ознаки. У клітинах шкіри організмів, гомозиготних по рецесивним аллелям всіх генів, міститься мінімальна кількість пігменту.
У деяких випадках домінантні і рецесивні алелі полигенов можуть забезпечувати розвиток різних варіантів ознак.
Наприклад, у рослини пастушої сумки два гена однаково впливають на визначення форми стручочки. Їх домінантні аллели утворюють одну, а рецесивні - іншу форму стручочки. При схрещуванні двох дигетерозигот за цими генами (рис. 7.16) в потомстві спостерігається розщеплення 15: 1, де 15/16 нащадків мають від 1 до 4 домінантних алелів, а 1/16 не має домінантних алелів в генотипі.
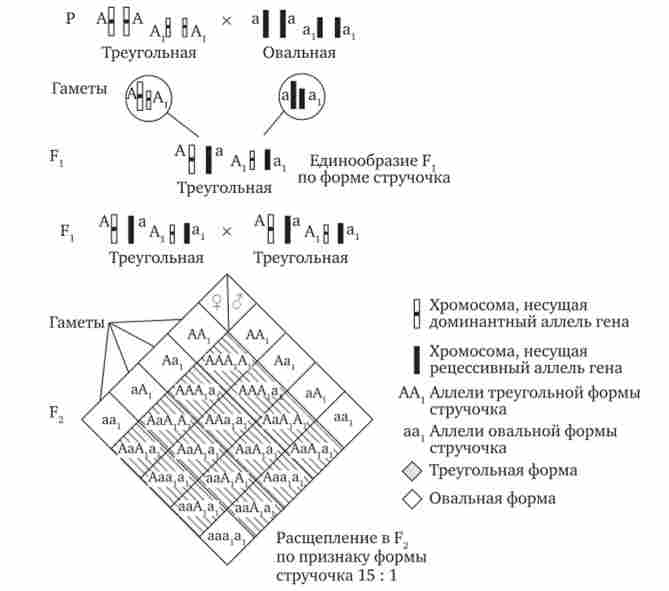
Мал. 7.16. Полімерне успадкування форми стручочки у грициків
Спадкування при комплементарном взаємодії генів.
Якщо складний ознака формується в результаті взаємодоповнююче дії певних алелей неалельних генів, то, очевидно, він буде з'являтися лише у тих організмів, які мають в генотипі саме таку комбінацію алелей.
Наприклад, присутність в генотипі домінантних алелів обох неалельних генів забезпечує розвиток складного ознаки, чого не відбувається при відсутності одного з них в домінантному стані. В цьому випадку при схрещуванні двох дігетерозіготних організмів, що мають цей показник, лише у певній частині потомства (9/16) буде формуватися така ознака, а у решти (7/16) він не розвинеться (рис. 7.17).
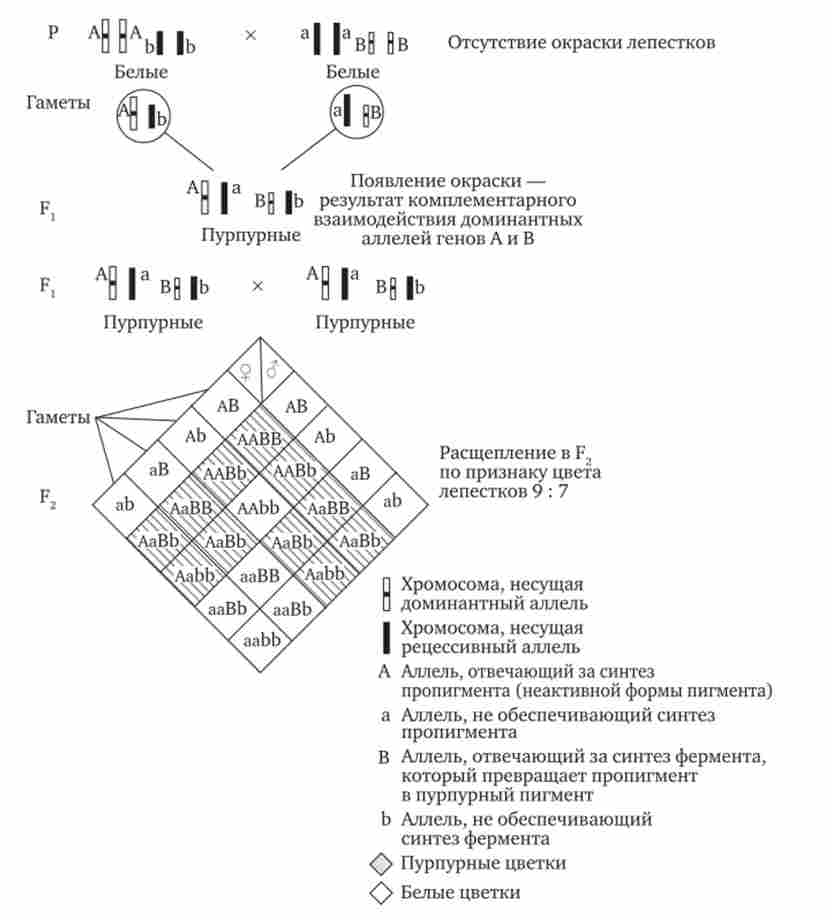
Мал. 7.17. Комплементарна взаємодія генів (успадкування ознаки забарвлення пелюсток у запашного горошку)
Можлива також ситуація, коли кожен з неалельних генів під час відсутності домінантного алеля іншого забезпечує розвиток певного варіанту ознаки, а разом вони формують новий його варіант (рис. 7.18). Тоді розщеплення в потомстві двох дигетерозигот буде відповідати розщепленню при незалежному успадкуванні ознак (9: 3: 3: 1).
У людини два гена, що детермінують відкладення в волоссі чорного і червоного пігментів; при певних поєднаннях їх алелей забезпечують поява нової ознаки - особливого блиску волосся.

Мал. 7.18. Комплементарна взаємодія генів (успадкування ореховіднимі форми гребеня у курей).
I - A? Bb - розовідний; II - Аав? - гороховідная; III - А? В? - ореховіднимі; IV - aabb - листоподібна
При Епістаз один з генів (В) виражається фенотипично лише при відсутності в генотипі певного алеля іншого гена (А). В його присутності дію гена В не виявляється. У строгому сенсі слова, цей вид взаємодії неалельних генів може бути розглянутий як варіант взаємодоповнююче дії певних алелей цих генів, коли один з них здатний забезпечити розвиток ознаки, але лише в присутності певного алеля іншого гена. У цій ситуації фенотип організму залежить від конкретного поєднання алелей неалельних генів в їх генотипах і розщеплення за фенотипом у потомстві двох дигетерозигот за цими генами може бути різним.
при домінантному епістаз, коли домінантний аллель одного гена (А) перешкоджає прояву алелей іншого гена (В або Ь), розщеплення в потомстві залежить від їх фенотипова значення і може виражатися співвідношеннями 12: 3: 1 або 13: 3 (рис. 7.19). При рецесивним Епістаз ген, що визначає якусь ознаку (В), не проявляється у гомозигот за рецесивним алелем іншого гена (аа). Розщеплення в потомстві двох дигетерозигот за такими генам буде відповідати співвідношенню 9: 3: 4 (рис. 7.20). Неможливість формування ознаки при рецессивном Епістаз розцінюють також як прояв нереалізованого комплементарного взаємодії, яке виникає між домінантним алелем Епістатичний гена і алелями гена, що визначає цю ознаку.
З цієї точки зору може бути розглянуто «бомбейський феномен» у людини, при якому у організмів - носіїв домінантного алеля гена, що визначає групу крові за системою АВО (1А або 1в), Фенотипично ці алелі не проявляються і формується I група крові (див. Рис. 3.82). Відсутність фенотипічного прояву домінантних алелей гена I пов'язують з гомозиготностью деяких організмів по рецесивним аллели гена Н (hh), що перешкоджає формуванню антигенів на поверхні еритроцитів. У шлюбі дигетерозигот по генам Н і I (НИА1В) 1/4 потомства буде мати фенотипічно I групу крові в зв'язку з їх гомозиготностью по рецесивним аллели гена Н - hh.
Розглянуті вище розщеплення за фенотипом у потомстві від схрещування гетерозиготних батьків або аналізує схрещування як при моногенному типі успадкування ознак, так і в разі взаємодії неалельних генів носять імовірнісний характер. Такі розщеплення спостерігаються лише в тому випадку, якщо реалізуються всі можливі зустрічі різноманітних
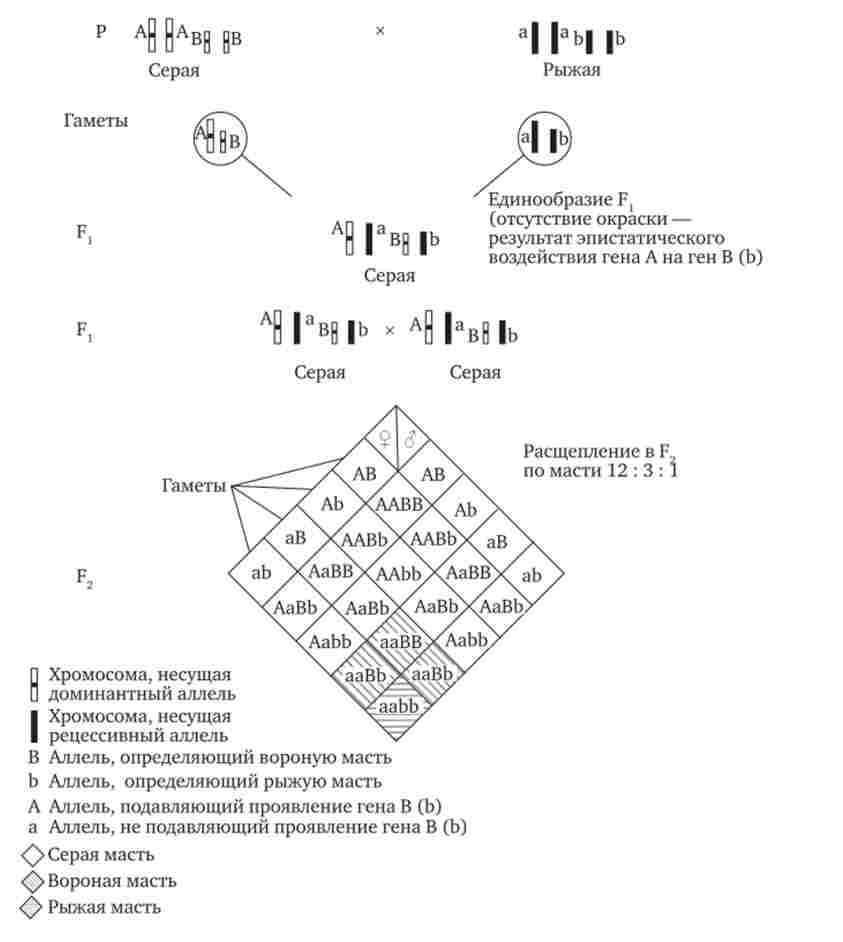
Мал. 7.19. Епістатичний взаємодія генів. Домінантний епістаз (успадкування масті у коней) гамет при заплідненні і всі нащадки виявляються життєздатними. Виявлення близьких розщеплення ймовірно при аналізі великої кількості нащадків, коли випадкові події не здатні змінити характер розщеплення. Г. Мендель, який розробив прийоми гибридологічного аналізу, вперше застосував статистичний підхід до оцінки отриманих результатів. Він аналізував велике число нащадків, тому розщеплення за фенотипом, що спостерігаються їм в дослідах, виявилися близькими до розрахункових,

Мал. 7.20. Епістатичний взаємодія генів. Рецесивний епістаз (успадкування пігментації шерсті у мишей) які виходять при обліку всіх типів гамет, утворених в мей- озе, і їх зустрічей при заплідненні.