| Головна |
| «« | ЗМІСТ | »» |
|---|
Спочатку в молекулярній генетиці вивчали гени головним чином найпростіших організмів: бактерій і їх паразитів - вірусів. У другій половині 1970-х рр. у біологів з'явилася техніка, яка дозволила їм вивчати структуру і функції генів у більш складних організмів, таких як ссавці, птахи, рослини та ін. Саме тоді було виявлено, що гени цих організмів відрізняються від генів бактерій. Виявилося, що це не цілісні освіти, а складаються з «частин і шматочків», що було несподіванкою для генетиків. Іншими словами, якщо уявити собі гени у вигляді тексту, що містить генетичну інформацію, то у бактерій він безперервний, а у вищих організмів розчленований безліччю вставок з інших текстів.
Отже, при вивченні структурних генів у еукаріот було встановлено, що багато хто з них, крім відрізків ДНК, що кодують амінокислоти в по- ліпептідной ланцюга, включають в свій склад «зайві», «мовчазні» ділянки ДНК, які не кодують амінокислоти. За пропозицією В. Джильберта вони були названі интронами, а ділянки генів, що кодують амінокислоти, - екзонами. Інтрони можуть бути також позначені як мднк (мовчить ДНК), а екзонів - як кДНК (кодує ДНК). Одночасне входження до складу генів екзонів і інтронів виявилося універсальної особливістю структурних генів еукаріот, якої позбавлені тільки гени, що кодують гістони, і псевдогени. Інтрони виявлені і в генах мітохондрій, а також в послідовності РНК і ДНК вірусів, що вражають клітини тварин, таких як аденовірус 2-го типу, SV40, вірус поліомієліту та ін.
Використання методів молекулярної генетики та електронномікроськопічеського аналізу дало можливість отримати більш повну інформацію про структуру та організацію багатьох структурних генів еукаріот. Виявилося, що число інтронів і їх положення в кожному з генів індивідуальні. У гені овальбуміна курки виявлено 7 интронов, а в гені колагену - 51.
Середня довжина інтронів становить кілька сотень пар нуклеотидів, а сумарна їх довжина зазвичай перевищує аналогічний показник екзонів у багато разів. Встановлено, що інтрони розташовані не по краях гена, а розподілені по всій його довжині. Іншими словами, гени вищих організмів складаються з кодують ділянок - екзонів, розділених некодуючими - нітроном. Визначення нуклеотиднихпослідовностей в інтрони на стику з екзонами виявило наявність динуклеотид АГ на З'-концс і динуклеотид ГТ на 5-кінці.
Освіта молекул мРНК у еукаріот характеризується двома особливостями. По-перше, спочатку транскрибується високомолекулярна пре-мРНК, в якій представлені копії як екзонів, так і інтронів. По-друге, оскільки трансляція вимагає наявності всього комплексу екзонів, при утворенні зрілих молекул мРНК діє механізм видалення всіх послідовностей, властивих Інтрони. При дозріванні молекул пре-мРНК ділянки, транскрибоване з інтронів, вирізаються, а що залишилися фрагменти, що містять Екзони, ковалентно зшиваються, утворюючи трансляційну систему гена, що складається тільки з екзонів, розташованих один за одним в потрібному порядку. Таким чином, генетична інформація набуває безперервну послідовність кодують нуклеотидів, подібну такої в класичному випадку з бактеріями. Процес зрощування екзонів в загальну нитка мРНК був названий сплайсингом (від англ, splice - зрощування кінців) (рис. 5.27).
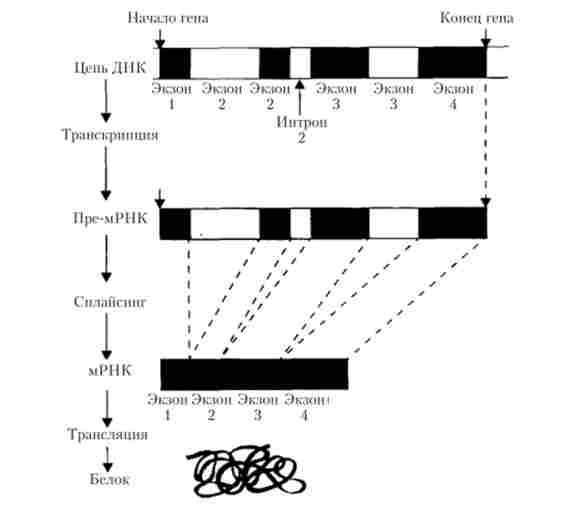
Мал. 5.27. Синтез білка з переривчастого гена
Таким чином, сплайсинг - це специфічний біохімічний процес, який призводить до вирізання нітронів і з'єднанню екзонів. Логічно було припустити, що в цьому процесі беруть участь ферменти, як і у всіх біологічних процесах. Тоді виникає питання про їх ідентифікації і про те, як їм вдається вирізати і з'єднувати послідовності нуклеотидних пар з такою точністю. Проблема розпізнавання меж интронов дуже важлива, адже якщо ферменти спрацюють неточно, то, з'єднавшись, Екзони будуть кодувати зовсім інший, неактивний білок (рис. 5.28). У таких випадках говорять про «зсуві рамки зчитування».
Нуклеотидну послідовність можна прочитати по одній з трьох рамок зчитування. При цьому кожен раз виходить нова послідовність амінокислот у синтезованої поліпегггідной ланцюга. У третьому варіанті зчитування з'являється терминирующего кодон, який перериває синтез білкової ланцюга.
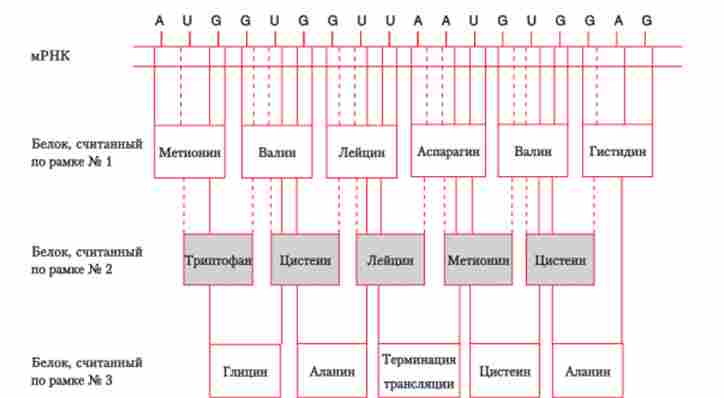
Мал. 5.28. Читання нуклеотидноїпослідовності
Отже, те, що сплайсинг пре-мРНК відбувається по межах інтронів і екзонів без зсуву рамки зчитування нуклеотидів, має особливе значення.