| Головна |
| «« | ЗМІСТ | »» |
|---|
В організмі пухка волокниста неоформлене сполучна тканина найпоширеніша. Вона розташовується близько епітеліальних тканин; в більшій чи меншій кількості супроводжує кровоносні, лімфатичні судини; входить до складу шкіри і слизових оболонок; у вигляді прошарків з судинами її виявляють у всіх тканинах і органах.
Пухка волокниста неоформлене сполучна тканина (рис. 31) складається з різноманітних клітин і міжклітинної речовини, що містить основне (аморфне) речовина і систему колагенових і еластичних волокон, розташованих неупорядоченно, тому тканина - неоформлене (див. Кол. Вкл., Рис. II) .
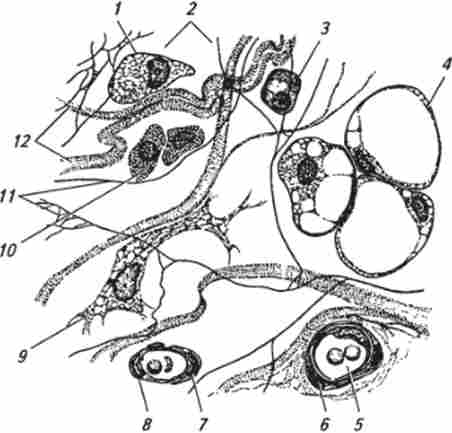
Мал. 31. Пухка волокниста неоформлене сполучна тканина:
I - макрофаг (гістіоціт); 2 - аморфне міжклітинний речовина; 3 - плазмоііт; 4 - жирові клітини; 5 - клітини крові в кровоносній судині; 6 - гладком'язових клітина; 7- адвентициальная клітина; 8 - ендотеліальна клітина; 9 - фібробласт; 10 - огрядні клітини (лабро- ціти); 11 - еластичні волокна; 12 - колагенові волокна
Поширеність, різноманітність і велика кількість клітинних елементів і міжклітинної речовини пухкої волокнистої неоформленої сполучної тканини забезпечують наступні функції:
трофічну - обмінні процеси, регуляція харчування клітин;
захисну - участь в імунних реакціях;
пластичну - відновлювальні процеси при тканинному пошкодженні;
опорну - освіту строми органів, зв'язування тканин органів між собою.
Клітини пухкої волокнистої неоформленої сполучної тканини в сукупності становлять собою єдиний дифузно розосереджений апарат, що знаходиться в нерозривному зв'язку з клітинами крові і лімфоїдної системою організму.
У пухкоїволокнистої неоформленої сполучної тканини є різноманітні високоспеціалізовані клітини: адвентіціальние, фібробласти, макрофаги, огрядні, плазматичні, жирові, пігментні.
Адвентіціальние клітини (від лат. Adventicus - сторонній, блукаючий) найменш диференційовані, багато в чому нагадують клітини мезенхіми, мають витягнуту зірчасті форму, часто з довгими відростками. Ці клітини розташовуються уздовж зовнішньої поверхні капілярів. Так як адвентіціальние клітини - камбіальні, вони активно діляться митозом і диференціюються в фібробласти, міофібробласти і ліпоціти.
Фібробласти (від лат. Fibrin - білок, blastos - паросток) - продуценти білка, є постійними і найбільш численними клітинами. В ході зародкового розвитку фібробласти утворюються безпосередньо з клітин мезенхіми, в постембріональний період. Фібробласти формуються з адвентиціальних клітин при регенерації.
Фібробласти мають веретеноподібну форму, велике ядро, яке слабо забарвлюється, чітко видно 1 ... 2 ядерця. Цитоплазма периферії клітини дуже світла, тому контури клітин невиразні і зливаються з основною речовиною. Навколо ядра цитоплазма, навпаки, забарвлюється інтенсивно, за рахунок великої кількості гранулярних ендоплазматичної мережі.
Фібробласти - це рухливі клітини. В їх цитоплазмі розташовуються мікрофіламенти, що містять актин. Вони скорочуються, і відбувається рух. Рухова активність фібробластів посилюється при утворенні капсули із сполучної тканини при пораненнях.
У дорослих тварин фібробласти мають незначну кількість цитоплазми, такі високодиференційовані клітини називають фиброцитами.
Макрофаги (гістіоцити) - клітини, що володіють здатністю до фагоцитозу і накопичення зважених колоїдних речовин в цитоплазмі. Макрофаги беруть участь в загальних і місцевих захисних реакціях імунітету (від лат. Immunitas - звільнення від чого-небудь).
В умовах культивування макрофаги міцно прикріплюються до поверхні скла і набувають уплощенную форму.
Ядро макрофагів має чітко окреслені контури, містить грудочки хроматину, добре забарвлюється основними барвниками. Цитоплазма містить багато вакуолей, що свідчить про активну участь в обміні речовин. Контури цитоплазми чіткі, відростки у вигляді псевдоподий, тому клітина схожа на амебу.
Основоположником вчення про макрофагах є І. І. Мечников, який об'єднав ці клітини в єдину систему - макрофагальну. Пізніше патолог Ашофф запропонував називати її ретіку- лоендотеліальной системою.
Рухливі, активно фагоцитирующие вільні макрофаги утворюються з різних джерел: адвентиціальних клітин, моноцитів, лімфоцитів і стовбурових кровотворних клітин. Моноцити циркулюючої крові представляють рухливу популяцію щодо незрілих макрофагів на шляху від кісткового мозку в органи і тканини.
За класифікацією Всесвітньої організації охорони здоров'я (1972) макрофаги об'єднані в Систему фагоцитів - СМФ.
Макрофаги беруть участь у багатьох імунних реакціях: в розпізнаванні, переробки та подання антигену лімфоцитів, в міжклітинному взаємодії з лімфоцитами. Маючи здатність до направленого руху - хемотаксису, макрофаги мігрують у вогнище запалення, де стають домінуючими клітинами при хронічному запаленні. При цьому не тільки очищають вогнище від сторонніх часток і зруйнованих клітин, але і стимулюють в подальшому функціональну активність фібробластів.
При запаленні макрофаги приходять в стан роздратування, збільшуються в розмірах, пересуваються і перетворюються в структури, звані Полібласти.
При електронній мікроскопії на поверхні макрофагів видно довгі пластинчасті відростки, за допомогою яких при фагоцитозі вони захоплюють чужорідні частинки. Відростки, подібно псевдоподиями амеби, оточують чужорідну частку і зливаються на верхівці клітини. Захоплена частка виявляється всередині цитоплазми, оточується лизосомами і поступово перетравлюється.
Залежно від локалізації (печінка, легені, черевна порожнина і ін.) Макрофаги набувають деякі специфічні особливості будови і властивості. Однак всім макрофагів властиві деякі загальні ультраструктурні і цитохімічні ознаки. Завдяки наявності скоротних ниток - филаментов, забезпечують рухливість плазмолеми, клітини цієї системи здатні до утворення різних пристосувань, що полегшують захоплення частинок. Один з основних ультраструктурних ознак макрофагів - наявність в цитоплазмі численних ли зосом, які розщеплюють і переробляють захоплений матеріал.
Макрофаги беруть участь не тільки в фагоцитозі, а й представляють антиген для запуску ланцюга імунних реакцій, що призводять до формування імунітету. Основні функції, за допомогою яких макрофаги беруть участь в реакціях імунітету, можна розділити на чотири типи: хемотаксис; фагоцитоз; секреція біологічно активних сполук; переробка антигену (процесинг) і уявлення антигену імунокомпетентним клітинам, що формує імунну відповідь.
При наявності у вогнищі токсичних і стійких подразників (деякі мікроорганізми, хімічні речовини, малорозчинні речовини) за участю макрофагів формується гранульома, в якій шляхом злиття клітин можуть утворюватися гігантські багатоядерні клітини.
При проникненні чужорідних частинок безліч макрофагів щільно прилягають один до одного, з'єднуються відростками, утворюють інтердігітаціі (від лат. Inter - між, digitatio - пальцевидні освіти). Це добре помітно в культурі тканин: утворення гігантських багатоядерних клітин передує формування інтердігітацій. Іноді гігантська многоядерная клітина утворюється при багаторазовому розподілі амитозе ядра одного макрофаги.
Гладкі клітини (тканинні базофіли, лаброці- т и) виявлені у всіх ссавців, проте кількість у тварин різних видів і в сполучної тканини різних органів неоднакова. У деяких тварин, наприклад у морських свинок, тканинних базофілів багато, але мало базофілів крові: обернено пропорційна залежність між зазначеними клітинами свідчить про подібному біологічному значенні.
Значна кількість тканинних базофілів міститься в пухкої сполучної тканини поруч з епідермісом, епітелієм травного тракту, дихальних шляхів, матки. Часто огрядні клітини виявляють в пухкої сполучної тканини між часточками печінки, в нирках, ендокринних органах, молочній залозі і в інших органах.
За формою тканинні базофіли частіше овальні або кулясті, розміром від 10 до 25 мкм. Ядро розташоване центрально, завжди містить багато грудочок конденсованого хроматину. Електронно-мікроскопічними дослідженнями в цитоплазмі виявляють мітохондрії, рибосоми; ендоплазматична мережа і комплекс Гольджі розвинені слабко.
Найбільш характерна структурна особливість тканинних базової уста філів - наявність численних великих (0,3 ... 1 мкм) гранул, рівномірно заповнюють ббльшую частина обсягу цитоплазми. Гранули оточені мембраною і мають неоднакову електронну щільність.
Розташовуючись поблизу дрібних кровоносних судин, тканинні базофіли одні з перших реагують на проникнення антигенів. Характерне метахроматичні фарбування гранул тканинних базофілів обумовлено наявністю гепарину і гістаміну. Дегрануляція тканинних базофілів, викликана різними факторами, призводить до виділення гепарину - речовини, що перешкоджає згортанню крові. Навпаки, без руйнування цілісності гранул відбувається секреція гістаміну, що підвищує проникність капілярів, стимулює міграцію еозинофілів, активізацію макрофагів.
Крім того, в гранулах тканинних базофілів містяться найважливіші біологічні аміни - серотонін, дофамін, що мають різноманітне фармакологічна дія. Тканинні базофіли беруть участь в розвитку алергічних і анафілактичних реакцій.
На цитоплазматичної мембрані тканинних базофілів, так само як і у базофілів крові, знаходиться значна кількість імуноглобулінів класу Е (IgE). Зв'язування антигенів і утворення комплексу антиген-антитіло супроводжується дегрануляцією і виділенням з тканинних базофілів судинно-активних речовин, що обумовлюють появу місцевих і загальних реакцій.
Плазматичні клітини (плазмоцити) синтезують і виділяють основну масу імуноглобулінів - антитіл - білки, що утворюються у відповідь на впровадження антигену.
Плазматичні клітини зазвичай зустрічаються у власному шарі слизової оболонки кишечника, сальника, в сполучної тканини між часточками слинних, молочних залоз, в лімфатичних вузлах, кістковому мозку.
Клітини можуть бути округлої або овальної форми; на внутрішній стороні чітко окресленої ядерної оболонки радіально розташовуються грудочки хроматину. Цитоплазма через наявність великої кількості РНК різко базофильная, виняток становить лише невеликий ободок цитоплазми близько ядра - перінуклеар- ная зона. По периферії цитоплазми є численні дрібні вакуолі.
За походженням плазматичні клітини є кінцеві стадії розвитку В-лімфоцитів, які в ділянках свого розташування активізуються, інтенсивно розмножуються і перетворюються в плазмоцити.
Освіта плазмоцита з активізованого В-лімфоцити за участю Т-хелперів і макрофагів проходить наступні етапи: По-лімфоцит - »плазмобласти -> проплазмоціт -> плазмоціт. Перетворення зазначених клітинних форм відбувається протягом 24 год.
плазмобласти - велика клітинка з великим ядром, активно ділиться митозом. Проплазмоціт набагато менше, характеризується різко вираженою базофілія цитоплазми, в якій з'являється багато розширених цистерн гранулярних ендоплазматичної мережі.
Плазмоціт (зрілий плазмоціт) містить невелику, ексцентрично розташоване ядро, в якому грудочки хроматину розподілені як спиці колеса. Белоксинтезирующий механізм запрограмований на синтез антитіл певного різновиду. Кожна плазматична клітина певного клону здатна за 1 год синтезувати кілька тисяч молекул імуноглобулінів.
На заключній стадії розвитку плазмоцити містять потужний белоксинтезирующий апарат, за допомогою якого синтезують імуноглобуліни - антитіла. Синтезовані молекули надходять в просвіт цистерн, потім в комплекс Гольджі, звідти після приєднання вуглеводного компонента виділяються з клітини. Антитіла виділяються при руйнуванні клітини.
У цитоплазмі плазматичних клітин утворюються ацидофільні включення у вигляді гомогенних структур, інтенсивно забарвлюються еозином в рожевий колір. При цьому базофилия цитоплазми зникає, ядро фрагментируется; поступово округляючись, з ацидофільних структур утворюється ацидофільне тільце Русселя, розташоване в основній речовині пухкої волокнистої неоформленої сполучної тканини. Тельці Русселя складається з глобулінів і комплексу глобулінів з вуглеводами.
Жирові клітини (ліпоціти) розташовуються головним чином поблизу кровоносних судин, а також можуть формувати відкладення жирової тканини (textus adiposus). В ембріогенезі жирові клітини формуються з клітин мезенхіми. Попередниками для утворення нових жирових клітин в постембріональний період є адвентіціальние клітини, які супроводжують кровоносні капіляри.
Жирові клітини синтезують і накопичують в цитоплазмі запасні ліпіди, головним чином тригліцериди.
З жирових клітин утворені часточки різних розмірів. Між часточками знаходяться прошарки пухкої сполучної тканини, в яких проходять дрібні кровоносні судини і нервові волокна. Між жировими 'клітинами всередині часточок розташовуються окремі клітини сполучної тканини (фіброціти, тканинні базофіли), мережа тонких аргірофільних волокон і кровоносні капіляри.
Жирові речовини виявляють при використанні спеціальних барвників (судан III, Судан IV, чотириокис осмію). Ліпоціти мають перстневидного форму, велика частина обсягу клітини зайнята однієї великої краплею жиру, овальне ядро і цитоплазма знаходяться на периферії клітини (див. Кол. Вкп., Рис. III).
У багатьох частинах організму тварин утворюються значні скупчення жирових клітин, які називаються жировою тканиною. У зв'язку з особливостями природного забарвлення, будови та функції, а також розташування розрізняють у ссавців два різновиди жирових клітин і відповідно два типи жирової тканини: білу і буру.
Біла жирова тканина в організмі тварин різних видів і порід розподілена неоднаково. У значній кількості вона міститься в так званих жирових депо: підшкірна жирова клітковина, особливо розвинена у свиней, жирова тканина навколо нирок в брижі (приниркова клітковина), у деяких порід овець біля кореня хвоста (курдюк). У великої рогатої худоби м'ясних та м'ясо-молочних порід групи жирових клітин розташовуються в прошарках пухкої волокнистої неоформленої сполучної тканини скелетних м'язів. М'ясо, отримане від таких тварин, має найкращі смакові якості і називається «мармуровим».
Структурна одиниця білої жирової тканини - кулясті жирові клітини до 120 мкм в діаметрі. При розвитку клітин жирові включення в цитоплазмі з'являються спочатку у вигляді дрібних розсіяних крапель, пізніше зливаються в одну велику краплю.
Загальна кількість білої жирової тканини в організмі тварин різних видів, порід, статі, віку, вгодованості коливається від 1 до 30% живої маси тіла. Запасні жири - найбільш висококалорійні речовини, при їх окисленні в організмі звільняється велика кількість енергії (1 г жиру = 39 кДж).
Підшкірна жирова клітковина має велике значення для захисту організму від механічних пошкоджень, захищає від втрат тепла. Жирова тканина уздовж нервово-судинних пучків забезпечує відносну ізоляцію, захист і обмеження рухливості. Скупчення жирових клітин в поєднанні з пучками колагенових волокон в шкірі підошов і лап створюють амортизацію при русі. Жирова тканина служить депо води. Освіта води - важлива особливість обміну жирів у тварин, що мешкають в посушливих районах (верблюди).
При голодуванні організм використовує перш за все запасні жири з клітин жирових депо, в яких зменшуються і зникають жирові включення. Жирова тканина очної орбіти, епікарда, лап зберігається навіть при сильному виснаженні.
Колір жирової тканини залежить від виду, породи і типу годівлі тварин. У більшості тварин, за винятком свиней і кіз, в жирі міститься пігмент каротин, що надає жовтий колір жирової тканини. У великої рогатої худоби жирова тканина перикарда містить багато колагенових волокон. Нирковим жиром називають жирову тканину, навколишнє сечоводи.
В області спини жирова тканина свиней містить м'язову тканину, а також нерідко волосяні цибулини (щетину) і волосяні сумки. В області очеревини є скупчення жирової тканини - так званий брижових, або мезентеріальний, жир, де міститься дуже велика кількість лімфатичних вузлів, що прискорюють окисні процеси і псування жиру. У брижове жирі часто зустрічаються кровоносні судини, наприклад у свиней більше артерій, а у великої рогатої худоби - більше вен.
Внутрішнє сало являє собою жирову тканину, розташовану під очеревиною. Воно містить велику кількість волокон, розташованих в косому і перпендикулярному напрямках. Іноді в жировій тканині свиней виявляють пігментні зерна, в таких випадках виявляються коричневі або чорні плями.
Бура жирова тканина в значній кількості є у гризунів і тварин, що впадають в зимову сплячку, а також у новонароджених тварин інших видів. Ця тканина розташована переважно під шкірою між лопатками, в шийній області, в середостінні і вздовж аорти. Бура жирова тканина складається з відносно дрібних клітин, дуже щільно прилягають один до одного, нагадуючи зовні железистую тканину. До клітинам підходять численні нервові волокна, обплетені густою мережею кровоносних капілярів.
Для клітин бурої жирової тканини характерні центрально розташовані ядра і наявність в цитоплазмі дрібних жирових крапель, які не зливаються у велику краплю. У цитоплазмі між жировими краплями розташовані гранули глікогену і численні мітохондрії, пофарбовані білки системи транспортних електронів -? цитохроми, які надають бурий колір цієї тканини.
У клітинах бурої жирової тканини інтенсивно відбуваються окислювальні процеси з виділенням значної кількості енергії. Однак більша частина утворюється енергії витрачається не на синтез молекул АТФ, а на теплоутворення. Така властивість липоцитов бурою тканини є важливим для регуляції температури у новонароджених тварин і для зігрівання тварин після пробудження від зимової сплячки.
Пігментні клітини (пігментоціти), як правило, мають відростки, в цитоплазмі дуже багато темно-коричневих або чорних зерен пігменту з групи меланінів. У сполучної тканини шкіри нижчих хребетних: рептилій, амфібій, риб, міститься значна кількість пігментних клітин - хро- матофоров, що обумовлюють ту чи іншу забарвлення зовнішнього покриву і виконують захисну функцію. У ссавців пігментні клітини зосереджені переважно в тканинах очного яблука - склери, судинної і райдужної оболонках, а також в ресничном тілі.
Представлено двома компонентами: основним (аморфним) речовиною - безструктурним матриксом, що має студневидного консистенцію; колагеновими і еластичними волокнами, розташованими щодо пухко і безладно.
До складу основної речовини входять високомолекулярні кислі мукополісахариди: гіалуронова кислота, хондро- ітінсерная кислота, гепарин. Ці хімічні компоненти виділяються як з клітин, так і з плазми крові. Кількість цих речовин в різних ділянках сполучної тканини неоднакове. Близько капілярів і дрібних судин, в ділянках, що містять жирові прошарки, або в тканини, багатої ретикулярних клітинами, основної речовини мало, а на кордонах з епітелієм, навпаки, багато. У цих ділянках основна речовина разом з ретикулярними волокнами утворює прикордонні базальні мембрани, часто добре помітні.
Стан основного речовини може змінюватися, в залежності від цього змінюється і вид базальної мембрани. Якщо основна речовина рідке, то прикордонний шар має волокнисту структуру; якщо щільне, то контури волокон не виступають і мембрана виглядає гомогенною.
Основна речовина заповнює проміжки між клітинами, волокнами, судинами мікроциркуляторного русла. Безструктурне основна речовина на ранніх стадіях розвитку тканини в кількісному відношенні переважає над волокнами.
Основна речовина - гелеобразная маса, здатна в широких межах змінювати свою консистенцію, що істотно відбивається на його функціональні властивості. За хімічним складом це дуже лабільний комплекс, що складається з глікозаміноглі- канов, протеогліканів, глікопротеїдів, води і неорганічних солей. Найважливішим хімічним високополімерних речовиною в цьому комплексі є несульфатованих різновид гли- козаміногліканов - гіалуронова кислота. Нерозгалужені ланцюги молекул гіалуронової кислоти, утворюють численні вигини і формують своєрідну молекулярну мережу, в комірках і каналах якої знаходиться і циркулює тканинна рідина. Завдяки наявності таких молекулярних просторів в основній речовині є умови для пересування різних речовин від кровоносних капілярів і продуктів клітинного метаболізму в зворотному напрямку - до кровоносних і лімфатичних капілярах для подальшого виділення з організму.
Колагенові волокна мають вигляд смуговидних тяжів, орієнтованих в різних напрямках. Волокна не розгалужуються, вони Малорозтяжні, мають велику міцність на розрив (витримують до 6 кг на 1 мм2 поперечного перерізу), здатні об'єднуватися в пучки. При тривалому варінні колагенові волокна утворюють клей (від англ, kolla - клей).
Міцність колагенових волокон зумовлена тонкої структурної організацією. Кожне волокно складається з фібрил діаметром до 100 нм, розташованих паралельно один одному і занурених в межфібріллярное речовина, що містить протеїни, глікозаміноглікани і протеоглікани. Колагенові волокна неоднакові за ступенем своєї зрілості. У складі недавно утворених при запальної реакції волокон є значна кількість цементуючого полисахаридного речовини, яке здатне відновлювати срібло при обробці зрізів солями срібла. Тому молоді колагенові волокна часто називають аргірофільних, в зрілих волокнах кількість цієї речовини зменшується.
При електронній мікроскопії по довжині фібрили спостерігають характерну поперечну смугастість - чергування темних і світлих смуг з певним періодом повторюваності, а саме один темний і один світлий сегмент разом складають один період довжиною 64 ... 70 нм. Найвиразніше ця смугастість видно на негативно забарвлених препаратах колагенових фібрил. На позитивно забарвлених препаратах колагенових фібрил, крім основної темно-світлій періодичності, виявляють складний малюнок більш тонких електронно-щільних смужок, розділених вузькими проміжками шириною 3 ... 4 нм.
Фібрила складається з більш тонких протофібрілл з білка тро- поколлагена. Протофібрілли мають довжину 280 ... 300 нм і ширину 1,5 нм. Освіта фібрили - результат характерною угруповання мономерів в поздовжньому і поперечному напрямках.
Молекула тропоколагену має асиметричну структуру, де подібні послідовності амінокислот виявляються один навпроти одного, виникають вузькі вторинні Темна смуги. Кожна молекула тропоколагену є спіраль з трьох поліпептидних ланцюгів, утримуваних водневими зв'язками. Унікальна структура тропоколагену обумовлена високим вмістом гліцину, оксилізину і оксипроліну.
Еластичні волокна мають різну товщину (від 0,2 мкм в складі пухкої сполучної тканини до 15 мкм в зв'язках). На пофарбованих гематоксиліном і еозином плівкових препаратах сполучної тканини волокна мають вигляд виражених тонких розгалужених гомогенних ниток, які формують мережу. Для виборчого виявлення еластичних мереж використовують спеціальні барвники: орсеїном, резорцин-фуксин. На відміну від колагенових еластичні волокна не об'єднуються в пучки, мають малу міцністю, високою стійкістю до дії кислот і лугів, нагрівання, до гідролізу дії ферментів (за винятком еластази).
При електронній мікроскопії в будові еластичного волокна розрізняють більш прозору аморфну центральну частину, що складається з білка еластину, і периферичну, в якій міститься велика кількість електронно-щільних микрофибрилл глікопротеідной природи, що мають форму трубочок діаметром близько 10 нм.
Освіта еластичних волокон в сполучної тканини обумовлено синтетичною і секреторною функціями фібробластів. Вважається, що спочатку в безпосередній близькості від фібробластів утворюється каркас з микрофибрилл, а потім посилюється утворення аморфної частини з попередника еластину - проеластіна. Під впливом ферментів молекули проеластіна коротшають і перетворюються на невеликі, майже сферичні молекули тропоеластіна. При утворенні еластину молекули тропоеластіна з'єднуються між собою за допомогою десмозіна і ізодесмозіна, відсутніх в інших білках. Крім того, в еластині немає оксилізину і полярних бічних ланцюгів, що обумовлює високу стійкість еластичних волокон.