| Головна |
| «« | ЗМІСТ | »» |
|---|
Це більш складний процес, так як транскрипція і трансляція відбуваються в різних компартментах і забезпечуються великою кількістю відповідних структур.
На рівні транскрипції регуляторні механізми у прокаріотів і еукаріот мають ряд обшіх рис. Розглянемо деякі відмінні риси. Для клітин еукаріот характерна ампліфікація генів і їх перебудова. Обидва механізми забезпечують різке збільшення копій тих чи інших білків, необхідних для реалізації клітинного метаболізму.
Відомо, що в клітинах еукаріот ДНК, поєднана з білками (гистонами), упакована в нуклеосоми (гл. 14). У цьому стані транскрипція неможлива, і для експресії генів необхідно деблокування транскріптона. Отже, утворення і руйнування нуклеосом є важливим фактором регуляції еукаріотичних генів. Яким же чином відбувається деблокування транскріптона?
Фосфорилювання гістонів. В результаті дії білкових гормонів відбувається опосередковане фосфорилювання ядерних білків - гістонів і руйнування нуклеосом. Матриця при цьому стає доступною для основних факторів ініціації транскрипції, і починається синтез РНК. При пре- фарбуванні дії гормонів нуклеосоми відновлюються.
Ацетилювання і деацетилювання гістонів. Це важливий фактор регуляції генної активності. Виявилося, що фермент гистон-ацетілаза асоційована з фактором ТАФ (гл. 28). Ацетилювання проходить по термінальному залишку лізину в поліпептидного ланцюга гистона. В результаті ацетилювання позитивний заряд білка зменшується і спорідненість гистона до негативно зарядженої ДНК знижується. Це може привести до руйнування нуклеосом і деблокування транскріптона. Деацетилювання гістонів призводить до протилежного ефекту. Специфічні ацетілаза і деацеті- лазу асоційовані з білками ініціації транскрипції.
Регуляторними елементами є білки инициаторного комплексу, описані в гл. 28. На додаток можна відзначити особливі нуклеотидні послідовності, що сприяють інтенсифікації транскрипції, так звані енхансери. Характерна особливість цих структур полягає в тому, що вони впливають на швидкість транскрипції незалежно від локалізації в оперон. Білки, які взаємодіють з енхансером, називаються енхансернимі елементами, розташованими на відстані 1000-2000 пар основ від регіону промотора. Ці білкові чинники здатні впливати на ініціацію транскрипції завдяки освіті ДНК-петлі, що призводить до просторового зближення енхансерних елементів і, наприклад, білків ТАТА (рис. 29.7).
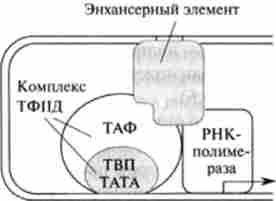
Мал. 29.7. Взаємодія енхан- серіого елемента з білками инициаторного комплексу
Дуже істотним чинником регуляції транскрипції є процесинг РНК. Освіта зрілих мРНК залежить від швидкостей кепірованія, освіти Поліани, а також швидкості стшйсінга. Для поліцістронной мРНК певний регуляторний значення має альтернативний сплайсинг (гл. 30).
Крім білків инициаторного комплексу, на швидкість транскрипції істотно впливають ДНК-зв'язуючі білки. З декількох сімейств найбільш відомі білки типу: цинкові пальці, спіраль-виток-спіраль і гомеодоменние білки. Специфічне зв'язування цих білків з ДНК відбувається в результаті взаємодії бічних радикалів амінокислотних залишків білка з основами ДНК.
Цинкові пальці представляють собою серію повторюваних доменів (від двох до дев'яти), що мають форму пальця. У центрі координації кожного домена знаходиться цинк. В одних випадках цинк з'єднаний з чотирма залишками цистеїну, в інших - з двома цистеїну і двома Гістидину.
На кінцевий результат - синтез білка - впливає також швидкість транспорту РНК в цитоплазму. У цитоплазмі мРНК, взаємодіючи з певними білками, утворює інформосому - своєрідне депо, з якого мРНК звільняється в міру потреби для синтезу білка. Швидкість звільнення мРНК також є фактором регуляції білкового синтезу.
Швидкість синтезу білка безпосередньо залежить від кількості мРНК, яке визначається часом її «напівжиття» або стабільністю in vivo. Таким чином, фактори, що впливають на стабільність мРНК, є регуляторами експресії генів і, як наслідок, білкового синтезу. Однією зі структур, що визначають стабільність мРНК, є Поліани-послідовність на З'ОН-кінці. Фактори, що регулюють експресію генів і синтез білка, підсумовані в табл. 29.2.
Таблиця 29.2. Фактори, що впливають на регуляцію транскрипції еукаріот
|
число |
фактори |
число |
фактори |
|
Ампліфікація генів Перегрупування генів Білки інініаторного комплексу ДНК-связиваюшим білки |
|
Сплайсинг мРНК Стабільність мРНК Транспорт мРНК в цитоплазму |
Лимитирующей стадією процесу трансляції є її ініціація. Найбільш докладно описаний процес зміни швидкості ініціації трансляції в результаті фосфорилювання фактора ініціації IF2. Реакція каталізується ферментом 1Р2-кінази, причому приєднання фосфатної групи інактивує фактор ініціації. Цей феномен був вивчений на прикладі синтезу гемоглобіну в ретикулоцитах. Спочатку було встановлено, що глобин синтезується тільки в присутності гема. Потім була вибудувана вся система регуляції синтезу глобіну. Виявилося, що активація 1Р2-кінази відбувається за рахунок її фосфорилювання цАМФ-залежної протеїнкінази. Взаємодія цієї протеїнкінази з цАМФ і її активацію блокує гем, виконуючи тим самим негативний контроль синтезу гемоглобіну.
Регуляція синтезу білка здійснюється також на стадії процесингу білка. Модифікації новосинтезованих поліпептидів здійснюються за допомогою відповідних ферментів, активність яких, в свою чергу, знаходиться під генетичним контролем. До цих модифікацій належать метилювання, фосфорилювання, гликозилирование, а також обмежений протеоліз.