| Головна |
| «« | ЗМІСТ | »» |
|---|
Середній мозок, так само як довгастий мозок і варолиев міст, відноситься до стовбурових структур. Він являє собою відносно невелику частину мозкового стовбура. Через середній мозок проходять все висхідні шляхи, що передають імпульси до таламуса, великих півкуль і мозочка, а також спадні шляху, які проводять імпульси до довгастого і спинного мозку. У середньому мозку, так само як і в довгастому, розташовані нейрони ретикулярної формації.
Анатомічно середній мозок складається з двох основних частин: спинного відділу, який являє собою кришку мозку, або четверохолмие, і базального, який представлений його ніжками. Кожна з ніжок, в свою чергу, складається з покришки, чорної субстанції і підстави. Покришка - це продовження дорсальній частині моста, в якій знаходяться червоне ядро і ядро блокового і окорухового нервів (рис. 13.14).
Мал. 13.14. Розріз через середній мозок на рівні ядер окорухового нерва:
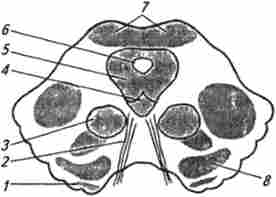
/ -Ножкі мозку; 2 окоруховий нерв; 3 червоне ядро; 4 - ядро окорухового нерва; 5 - центральна сіра речовина; б-водопровід мозку (Сільвією водопровід); 7- переднє двухолмие; 8- чорна субстанція
Рефлекторні функції ядер середнього м о з - г а. Передні горби четверохолмия є первинними зоровими центрами, за участю яких здійснюються деякі рефлекси у відповідь на світлові подразнення. До таких рефлексам ставляться зорові орієнтовні рефлекси, які проявляються в тому, що тварина, позбавлене великих півкуль, але зі збереженим середнім мозком, реагує на світлове роздратування рухом очей і поворотом голови. Рефлекторні рухи очей відбуваються завдяки надходженню до очних м'язів імпульсів від ядер окорухового і блокового нервів. Передні горби четверохолмия беруть участь в здійсненні зрачкового рефлексу, акомодації очі, а також в конвергенції - зведенні зорових осей.
Задні горби четверохолмия являють собою первинні слухові центри. За їх участі здійснюються орієнтовні слухові рефлекси. При орієнтовних рефлексах у тварин з цілим середнім мозком спостерігаються і вегетативні рефлекси: зміна ритму серцевої діяльності, артеріального тиску.
ядра четверохолмия забезпечують сторожовий рефлекс, значення якого полягає в підготовці реакції організму на раптове роздратування. При цьому відбувається перерозподіл м'язового тонусу: посилення тонусу згиначів, що сприяє швидкій реакції тварини.
В ніжках мозку йдуть провідні шляхи як від нижчих центрів в вищі центри головного мозку, так і від вищих центрів в спинний і довгастий мозок. Ці ж провідні шляхи утворюють ніжки мозочка і пучки волокон між скупченнями нервових клітин.
Чорна субстанція відповідає за координацію актів ковтання і жування, а також бере участь в регуляції пластичного тонусу; має значення при виконанні пальцями дрібних рухів, що вимагають великої точності. Тому чорна субстанція краще розвинена у приматів.
червоне ядро - велике скупчення сірої речовини середнього мозку, складається з великих і дрібних клітин. Воно пов'язане провідними шляхами з мозочком і ядрами смугастого тіла, а також зі спинним мозком (руброспінальний шлях). Червоні ядра з навколишнім їх ретикулярної формацією, безсумнівно, беруть участь в регуляції м'язового тонусу. Можна порівняти поведінку деце- ребрірованного того, що має зі спинним і довгастим мозком зберігся зв'язок лише за допомогою варолиева моста, мозочка і нижньої ділянки середнього мозку без області червоних ядер, з поведінкою так званого таламичного того, що має видалені кора мозку і підкіркові вузли, але середній мозок з його червоними ядрами і зорові горби збережені і залишаються в зв'язку з довгастим мозком.
Тонічні рефлекси стовбура мозку. Найважливіша функція стовбура мозку, зокрема довгастого і середнього мозку, полягає в перерозподілі м'язового тонусу в залежності від положення тіла в просторі. Перерозподіл тонусу здійснюється рефлекторним шляхом і забезпечує збереження рівноваги тіла. Всю сукупність тонічних рефлексів голландський вчений Р. Магнус розділив на дві великі групи: статичні рефлекси; статокинетичні рефлекси.
Статичні рефлекси. Виражені при спокійному стоянні, лежанні і сидінні в різних позах, вони обумовлюють певне положення тіла в просторі. Такі рефлекси діляться на дві великі групи. Перша група забезпечує певне положення або позу тіла - це рефлекси положення, або по- зотонічні рефлекси. Друга ж сприяє поверненню тіла з неприродного положення в нормальне - це установчі, або випрямні, рефлекси.
Позотонічні рефлекси включають в себе лабіринтові і шийні. лабіринтові рефлекси, для здійснення яких досить наявності довгастого і спинного мозку, забезпечують фіксування кінцівок в певному положенні. В експерименті на шию децеребрированійі тваринного надягали гіпсову пов'язку, щоб голова зберігала постійне положення по відношенню до тулуба. При цьому не відбувається роздратування проприорецепторов шиї. Потім тварина обертали навколо осі, умовно проведеної через обидва скроні. У положенні, коли тварина була спиною вниз, тонус розгиначів ставав максимальним і кінцівки витягувалися, а якщо ж воно виявлялося спиною вгору, то тонус цих м'язів знижувався і кінцівки згиналися.
На тонус всіх м'язів крім лабіринтових впливають також шийні рефлекси. Вплив шийних рефлексів позначається в тому, що при поворотах голови по відношенню до тулуба справа наліво або зліва направо тонус розгиначів максимальний на тій стороні тіла, до якої звернено тім'я. Якщо ж повертати голову навколо осі, умовно проведеної через обидва слухових проходу, то при нахилі голови вниз тонус розгиначів збільшується в задніх і знижується в передніх кінцівках. При закиданні голови назад, навпаки, тонус розгиначів передніх кінцівок збільшується, а задні кінцівки згинаються.
Рефлекси з лабіринтів на шийні миіщи. Шийні і лабіринтові тонічні рефлекси здійснюються в зв'язку один з одним, так як імпульси з лабіринтів впливають на тонус шийних м'язів. Максимум тонусу шийних м'язів спостерігається при тих положеннях голови, коли імпульси з лабіринтів ведуть до рефлекторного посилення тонусу розгиначів кінцівок. Імпульси з кожного лабіринту впливають переважно на шийні м'язи протилежної сторони (тому при руйнуванні лабіринту голова тварини повернута в бік пошкодження).
Випрямні рефлекси здійснюються середнім мозком і, отже, відсутні у бульбарних тварин. У здійсненні цих рефлексів беруть участь рецептори лабіринтів, шийних м'язів і шкіри.
Якщо мезенцефально тварина покласти на бік, воно піднімає голову і встановлює її в природне положення тім'ям догори. Це відбувається в результаті порушення рецепторів вестибулярного апарату. Але навіть при руйнуванні вестибулярного апарату голова випрямляється в тому випадку, якщо тіло лежить на боці на який-небудь твердої поверхні. В цьому випадку у відповідь на одностороннє роздратування шкіри відбувається рефлекторне випрямлення голови. Якщо на це ж тварина покласти зверху дошку з невеликим вантажем, то відбувається симетричне двостороннє роздратування шкірних рецепторів і голова знову падає вниз. Якщо дошку прибрати, то голова рефлекторно знову піднімається.
Підйом голови - це лише перша фаза випрямних рефлексів. Друга фаза полягає в рефлекторному випрямленні тулуба, яке слід за підйомом голови. Цей рефлекс виникає від подразнення пропріорецепторов м'язів шиї і рецепторів шкіри тулуба.
Статокинетічеськие рефлекси. Починаються з одного рецептивного поля-з рецепторів півколових каналів внутрішнього вуха. Виникають при кутовому або лінійному прискоренні.
При обертанні тваринного спостерігається ністагм голови, т. Е. Повільний поворот в сторону, протилежну обертанню, і швидке повернення в початкове положення. Очі реагують на обертання аналогічним чином - очної ністагм. Здійснення статокинетичних рефлексів можливе лише за участі ядер середнього мозку.