| Головна |
| «« | ЗМІСТ | »» |
|---|
Довгастий мозок і варолиев міст разом складають задній мозок. Анатомічно і функціонально задній мозок пов'язаний зі спинним мозком, мозочком і великими півкулями головного мозку. У ньому замикаються дуги багатьох сложнокоордінірованним рухових процесів.
Сегментарна будова, чітко виражене в спинному мозку, стерто в довгастому. Сюди вступають аферентні волокна від слухових рецепторів, рецепторів порожнини рота, шкіри лицьової частини голови, серця, великих судин і легенів, слизових оболонок очей, ряду рецепторів шлунка, печінки, підшлункової залози, тонких кишок. Але в органах грудної та черевної порожнини є також рецептори, від яких аферентні волокна йдуть не в довгастий, а в спинний мозок. Еферентні волокна нейронів довгастого мозку іннервують всі м'язи обличчя, серце, бронхи, гортань, стравохід, шлунок, підшлункову залозу, печінку, кишечник. Іннервація внутрішніх органів здійснюється парасимпатичними волокнами спільно з симпатичними волокнами, що лежать в спинному мозку.
З області довгастого мозку і варолиева моста виходять такі черепно-мозкові нерви: трійчастий, відвідний, лицьова, слуховий, язикоглоткового, блукаючий, додатковий, підязиковий.
Довгастий мозок грає велику роль в регуляції м'язового тонусу. Скелетні м'язи знаходяться в постійному тонусі, за рахунок якого долається дію сили тяжіння і тіло зберігає рівновагу. Тонус м'язів має рефлекторну природу. Початок дуги цього рефлексу лежить в м'язах і сухожиллях.
Якщо головний мозок разом із середнім мозком і його червоними ядрами видалений, а довгастий мозок збережений і не відокремлений від спинного, то у ссавців розвивається сильне, годинами і цілодобово утримується напруга розгинальних м'язів (рис. 13.13).
Кінцівки такої тварини витягнуті, і зігнути їх можна лише з працею; шийні м'язи, що розгинають голову, напружені; хребет кілька вигнутий дугою догори.
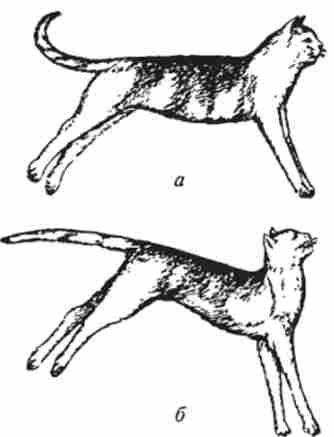
Таким чином, всі м'язи, напруга яких протистоїть дії сили тяжіння, виявляються після відділення довгастого мозку від середнього в стані тривалого тонічного напруги. це
Мал. 13.13. Деіерсбраціонная ригідність:
а-кішка з вимкненими великими півкулями (шляхом перев'язки судин), б- то ж при одночасному виключенні мозочка різке посилення тонусу розгинальних м'язів носить назву децеребрационной ригідності (Децеребрація - видалення мозку вище довгастого). Тонус розгиначів при децеребрационной ригідності перевищує тонус згиначів, але тривале напруження згиначів сприяє тому, що кінцівки стають ніби твердими опорами і можуть утримувати вагу тіла.
Децеребрационная ригідність не настає, якщо середній мозок тварини пов'язаний з довгастим мозком, а також якщо довгастий відділений від спинного мозку. Звідси випливає, що в довгастому мозку лежать нервові клітини, які можуть призводити до порушення спинальні нейрони, що іннервують розгиначі. Якщо довгастий мозок пов'язаний із середнім мозком, ригідність також не настає, т. Е. Від середнього мозку виходять впливу, які пригнічують стимуляцію розгинальних рефлексів з боку довгастого мозку.
Заключний етап у виникненні непомірного тонусу пов'язаний з рефлекторною діяльністю ядра вестибулярного нерва. Волокна від рецепторів вестибулярного апарату в довгастому мозку контактують з масою клітин, що утворюють так зване вестибулярное ядро. Оперативне видалення вестибулярного ядра і особливо його частини, відомої як ядро Дейтерса, різко послаблює децеребрационной ригідність. Руйнування разом з ядром Дейтерса формації довгастого мозку абсолютно її припиняє.
Для розвитку децеребрационной ригідності необхідне надходження в ЦНС імпульсів від рецепторів розгинальних м'язів. Ін'єкція в ці м'язи розчинуновокаїну або перерізання відповідних задніх корінців знімає тонус м'язів-раз-гібателей. Отже, децеребрационной ригідність треба розглядати як результат посилення рефлексів, здійснюваних спільно спинним і довгастим мозком у відповідь на імпульси, що надходять як від рецепторів м'язів-розгиначів, розтягуваних під дією сили тяжіння, так і від рецепторів лабіринту. Ці рефлекси в нормі завжди врівноважуються впливами, рефлекторно виникають в середньому мозку, мозочку, а у вищих ссавців головним чином в корі великих півкуль.
Роль довгастого мозку в здійсненні децеребрационной ригідності обумовлена постійним надходженням імпульсів від найважливіших рецепторовдихательной і кровоносної систем, а також з лабіринтів, внаслідок цього він постійно впливає на рефлекторні центри спинного мозку, підвищуючи їх збудливість. При відділенні спинного мозку від довгастого збудливість нервового центру ніколи не досягає того рівня, при якій тонус м'язів-розгиначів у відповідь на імпульси від їх рецепторів рефлекторно посилювався б до ступеня, що характеризує децеребрационной ригідність.
Децеребрационная ригідність - це стан, звичайно, абсолютно нефізіологічна, але по ній можна судити про взаємини між різними утвореннями ЦНС. Треба зауважити, що механізм цього процесу ще не зовсім ясний. Цей стан не завжди настає після децеребрации, іноді воно виражено дуже різко, а іноді майже не проявляється.
Довгастий мозок бере участь в регуляції провідникових рефлексів і в здійсненні таких актів, як ходьба і стояння, для яких необхідна інтегративна діяльність центральної нервової системи. Бульбарний тварина, у якого збережені лише спинний і довгастий мозок, здатне до більш складним рефлекторним процесам, ніж спинальне, так як всі основні функції в більш повній мірі об'єднані і скоординовані.
Варолиев міст є центральною частиною заднього мозку. Він має вигляд товстого білого валу. Сіра речовина утворюють ядра черепно-мозкових нервів (V-VIII пари), ядра ретикулярної формації і власні ядра мосту.
Біла речовина складається з поздовжніх і поперечних пучків нервових волокон. Поздовжні волокна йдуть по нижній поверхні моста в довгастий мозок, утворюючи в ньому піраміди. Зверху вони складають висхідні і низхідні шляхи двосторонніх зв'язків моста. Поперечні волокна направляються в ніжки мозочка.
Чутливі волокна черепно-мозкових нервів проводять сигнали від смакових рецепторів язика, рецепторів шкіри і м'язів голови, очей і зубів. Еферентні волокна забезпечують міміку. Таким чином, рефлекторна діяльність варолиева моста доповнює захисні і харчові рефлекси, здійснювані довгастим мозком.